

撰文|摩西
編輯|南風
正文共:16778字,預計閱讀時間:42分鐘
近幾十年來,MSCs在全球范圍內的實驗和臨床研究中涉及各種疾病,由于其獨特的生物學特性,顯示出優于常規治療的治療效果 [1,2,3,4,5]。它們從圍產期組織中分離出來,例如臍帶、臍帶血和胎盤,以及成人的多種生物組織,包括骨髓、脂肪組織、肌肉和肺 [6,7]。間充質干細胞作為一種多能成體干細胞,具有自我更新和多向分化為脂肪細胞、肌肉、軟骨細胞、成骨細胞和神經元細胞的潛力。[8,9]。此外,越來越多的證據表明,MSCs 通過高旁分泌活性發揮免疫調節、修復和再生作用[10,11,12](圖1)。更重要的是,MSCs 具有免疫特權,這意味著異基因MSCs移植不會引起炎癥反應,這主要是由于它們缺乏II類主要組織相容性復合物 (MHC-II) 和共刺激分子 [13,14]。這些特點使間充質干細胞成為繼造血干細胞之后細胞治療的理想種子細胞。
間充質干細胞的分離來源和作用機制。該圖說明了獲取MSCs的各種來源以及與治療效果相關的生物學特性。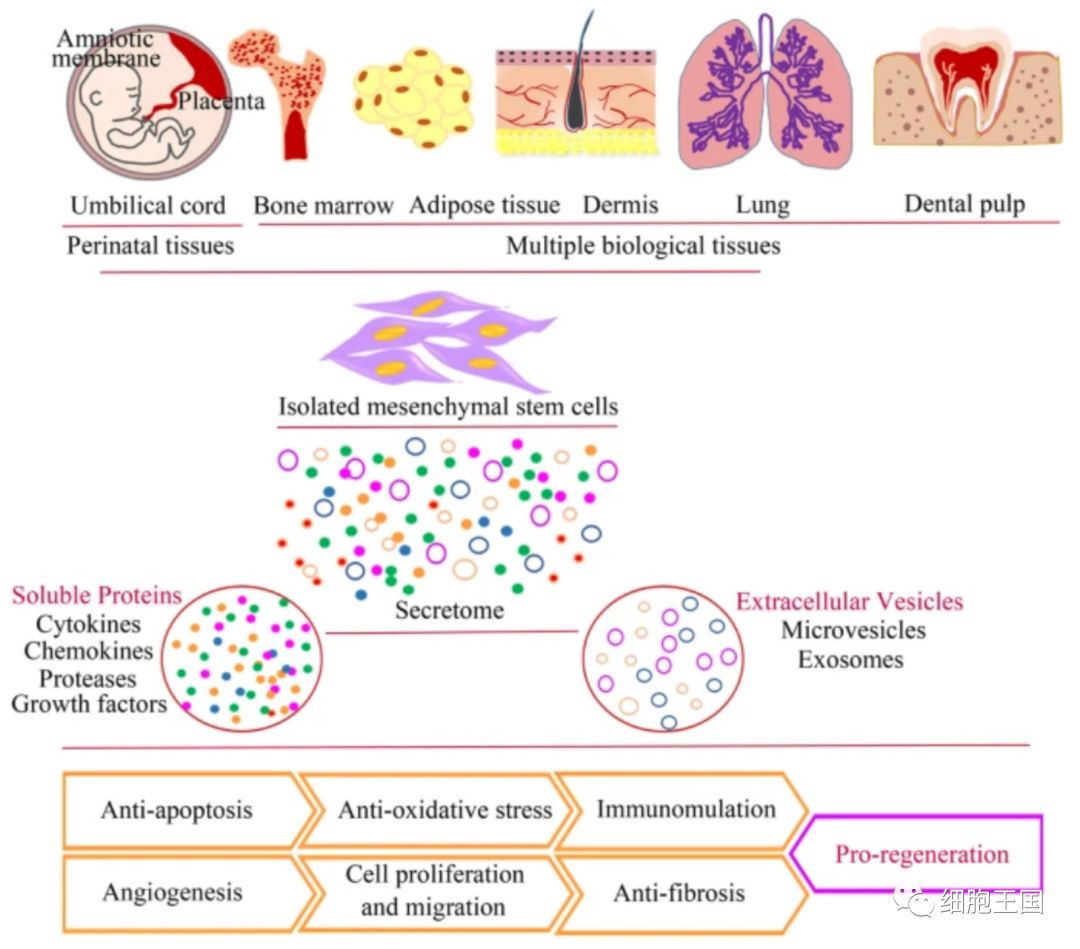

然而,來自不同個體的間充質干細胞的生物學效應具有較大差異,身體的內部環境(生病與否,年輕或年老)會影響分離出的干細胞的質量。例如,從肥胖和2型糖尿病 (T2D)個體中分離出的脂肪來源的MSC (ASC) 表現出功能缺陷,例如細胞凋亡增加、免疫抑制活性降低和干性喪失 [15,16,17]。來自老年供體的MSCs表現出增殖和分化能力受損、免疫調節抑制和生物活性分子分泌減少[18]。此外,一旦進入體內,MSCs可能會很快發生凋亡,因為它們暴露于惡劣的宿主微環境,包括缺氧、氧化應激以及慢性炎癥。
據報道,靜脈注射非健康個體來源的MSCs只有約28%存活超過1天[19],不到1%的細胞存活超過一周 [20,21]。即使原位移植,大多數MSCs也會在一周內失去其生物學活性 [22]。此外,大多數注入的MSCs被困在肺微血管而不是靶組織中 [23]。這些不利條件將導致各種問題,例如移植細胞的存活率低,MSCs的遷移和歸巢不良,從而限制了輸注細胞的功能。
為了達到預期的治療潛力,本文提出了幾種嘗試優化干細胞的策略,大致分為兩類,即基因改造和預激活(非基因改造)。
在基因改造方面,間充質干細胞會產生或過表達功能基因,使其能夠抵抗惡劣的微環境和細胞凋亡,增加遷移和歸巢,增強旁分泌作用。幾項研究表明,基因轉染的MSCs比野生型 MSCs具有更好的治療潛力 [25,26,27]。然而,安全性是轉基因MSCs未來臨床治療應用的最大障礙。據報道,病毒表達系統可以在宿主體內引發免疫和炎癥反應,而病毒在宿主基因組中的整合會帶來致瘤風險 [28,29]。此外,需要充分闡明基因工程MSCs的治療潛力和長期功能。因此,開發高效的非基因改造方法,在此統稱為預激活,是改善MSCs治療結果的一種替代和可操作的方法。
MSCs可以被預激活以實現所需的功能并逆轉它們的失活,因為它們可以識別微環境中的刺激并記住它們[3,30]。回顧目前的文獻,MSCs的預激活主要是基于MSCs存活的體內生理微環境將其在體外進行模擬,稱為“生理微環境模擬預激活”。或者MSCs的體外適應性調節是基于疾病的病理微環境,稱為“病理微環境模擬預激活”。評論文章的主要目標是提供涉及這兩種類型的預激活的具體方法(圖2)。

間充質干細胞的分離來源和作用機制。該圖說明了獲取MSCs的各種來源以及與治療效果相關的生物學特性。
o1
生理微環境模擬預激活
原代培養中的MSC數量有限。它需要在體外擴增以達到治療所需的細胞數量,過度的體外操作會導致衰老、干性下降和再生能力受損[23]。在體外保持MSCs的“年輕”非常重要。干細胞生活在組織的特定區域,稱為干細胞生態位。它是一個由細胞和非細胞成分組成的多維環境,用于操控干細胞增殖、決定干細胞命運和維持干細胞穩態。涉及許多必要線索的細胞和非細胞成分,即其他細胞(細胞-細胞相互作用)、物理因素(溫度、滲透壓、拉伸和電信號)、化學因素(PH、氧氣、營養物質、離子強度、代謝物細胞因子和趨化因子)和細胞外基質(組成、結構、拓撲結構和剛度)[31,32,33]。在這方面,在體外重建復雜的體內微環境將為MSC的生產提供有益的支撐,并保持其固有的生物學特性。迄今為止,對缺氧和三維 (3D) 培養進行了最深入的研究。
o2
低氧預激活
在體外培養ADCSs可能會面臨各種壓力,包括物理脫離、溫度變化和蛋白酶誘導的細胞損傷,從而導致細胞衰老。
在體外培養的MSCs通常暴露在平均氧分壓約為21% [34] 的環境中。然而,MSCs通常存在于低氧微環境中,體內生理氧濃度范圍為1%至11% [35,36]。多項研究表明,高氧濃度會導致培養的MSCs受到環境壓力,進而誘導DNA損傷和衰老 [37,38],并使其活性降低[39,40]。因此,低氧是MSCs生理微環境的重要組成部分。缺氧誘導因子 (HIF),尤其是 HIF-1,是細胞對缺氧反應的關鍵調節劑[41]。HIF-1是一種異二聚體,包含兩個亞基HIF-1α和HIF-1β。O2的缺乏使氧不穩定蛋白HIF-1α積累并易位進入細胞核,然后與HIF-1β 結合形成異二聚體,進一步與靶基因中的缺氧反應元件 (HRE) 結合。CBP/p300等激活劑,然后調節許多基因的轉錄 [42,43,44]。
低氧預激活對MSCs具有多種有益作用。例如,低氧培養環境保持MSCs的未分化狀態。一些報告表明,低氧張力增加了多能干細胞標志物(Oct4、Sox2 和 Nanog)在ASC中的表達,而不會改變它們的表面標志物和形態 [45, 46]。
此外,與常氧狀態(20% O2)相比,低氧促進了MSCs的增殖和存活,從而提高了擴增效率以獲得更多的ASCs產量[45,46,47,48]。并且在遷移試驗中,低氧啟動似乎促進了MSCs 在體外的動員[47,49] 并改善了MSCs 在體內的歸巢 [50,51]。李等人證明,低氧培養促進 了MSCs的增殖和遷移潛能[34]。Rosová 等人的結果,表明在低氧條件下培養的MSC增強了肝細胞生長因子 (HGF) 及其主要受體cMet的表達,而HGF/cMet是MSC遷移的主要信號[51]。另一項研究發現,低氧預激活通過增加鉀Kv2.1通道表達和FAK活性來促進骨髓間充質干細胞 (BMSCs) 的遷移 [50]。
此外,低氧啟動可以保護MSCs免受不利微環境的影響,并保持基因穩定性[37,52]。與常氧培養相比,MSCs暴露于低氧環境時,其壽命延長,可避免復制性衰老,并且表達較少的衰老相關β-半乳糖苷酶 [53]。低氧條件下的MSCs表現出基因穩定性,DNA損傷減少,染色體畸變減少 [37]。低氧培養使MSCs的分泌譜發生變化,旁分泌功能增強。例如,在缺氧條件下,MSCs 中促血管生成因子如 VEGF、HGF 和成纖維細胞生長因子(bFGF)的分泌增加。此外,MSCs的免疫抑制特性在低氧條件下得到改善,抗炎因子上調,抑制CD4和CD8 T淋巴細胞的增殖,促進Treg細胞的生成比暴露于正常氧氣的MSCs更有效[56,57]。
值得注意的是,大量實驗研究發現,低氧預激活的MSCs比未處理的MSCs顯示出更顯著的治療效果。在肺纖維化模型中,與低氧培養的MSCs相比,低氧預活化的MSCs可以明顯減輕肺水腫、減輕肺纖維化并改善肺功能 [58]。此外,將低氧MSCs移植到大鼠心肌梗塞模型中會改善受傷部位的血流和減少梗死面積 [59]。此外,與常氧MSCs治療的糖尿病小鼠相比,鏈脲佐菌素誘導的糖尿病小鼠接受低氧預活化的MSCs后,隨機血糖和空腹血糖顯著降低,口服葡萄糖耐量得到改善[40]。
因此,與常氧培養MSCs相比,低氧預處理的MSCs表現出更有利的特性并具有更好的治療潛力。此外,低氧預活化具有簡單、成本低、易于推廣、適合大規模細胞生產等優點。然而,在將低氧MSCs進一步應用于臨床之前,需要解決幾個問題。有必要優化低氧預激活中的氧濃度,因為MSC生活的生理生態位中的氧濃度因組織來源而異(骨髓中為1%至7%,脂肪組織中為10%至15%,脂肪組織中為1.5%至1.5%)[58]。此外,生物安全性是干細胞療法最受關注的問題,在臨床應用前應慎重確定。
o3 3D培養預激活
與傳統二維 (2D) 培養環境相比,3D培養系統模仿體內天然MSC微環境,增強了細胞-細胞相互作用或細胞-ECM相互作用,可顯著改善MSCs的生物學特性為,如增殖、免疫調節和定向分化[60]。MSCs的3D培養系統,包括多細胞球體、支架和水凝膠,越來越受到關注。
球體培養
球體培養是一種比較簡單的增強MSCs生物活性的方法 [61]。已經開發了各種方法來生成 MSC球體,例如懸滴技術、低附著方法以及強制聚集技術 [62]。隨著生物材料的發展,生產MSC的球體支架培養平臺已經發展起來。
球體培養系統可以通過增加干性和促進分化成不同的細胞譜系來提高MSCs的治療潛力[57,63]。Cheng等人的研究結果表明,基于殼聚糖薄膜的球體培養可以使間充質細胞系標記CD29、CD90和CD105的下調和多能性相關標記Sox2、Oct4、Nanog和SSEA-4的上調,使MSCs去分化成更原始的狀態[45,64]。此外,張等人采用微重力生物反應器生成 MSCs球體,并得出了類似的結論?[65]。球體衍生的MSCs改變了它們的分化偏好,可以轉分化為非間充質譜系細胞,如神經細胞和肝細胞 [64,65]。
球體培養系統可以通過提高增殖、遷移和歸巢效率來提高MSCs的治療潛力。與單層MSC 培養相比,球體來源的MSC表現出更高的增殖活性[66]。SDF-1/CXCR4信號通路在移植的MSCs的遷移和植入中起著至關重要的作用。球體培養系統生產的MSCs可恢復由2D培養體系引起的CXCR4表達缺失 [67]。此外,在球體衍生的MSCs中發現基質金屬蛋白酶MMP-9和MMP-13的表達增加,從而促進細胞通過基底膜侵入[66]。
球體培養系統可以通過促進治療因子(包括免疫調節和促血管生成細胞因子)的分泌來提高 MSCs的治療潛力。據巴托什等人報道,懸滴模型中的MSCs分泌更高水平的抗炎因子,如腫瘤壞死因子誘導基因6蛋白(TSG-6),更有效地抑制脂多糖(LPS)刺激的腫瘤巨噬細胞分泌壞死因子-α (TNF-α)?[68]。另外的幾項研究表明,其他抗炎細胞因子,如前列腺素E2 (PGE-2)、轉化生長因子-β1 (TGF-β1) 以及IL-6在球體系統培養的MSCs中也表現出更高的水平 [69,70,71,72] 。此外,由于VEGF、HGF、bFGF 和血管生成素 (ANG) 的分泌顯著增加,球體培養來源的MSCs 的促血管生成和促再生功能顯著增強 [67,73]。
球體培養優化了間充質干細胞的生物學特性,使其在體內具有更好的治療效果。巴托什等人證明,在酵母聚糖誘導的小鼠腹膜炎模型中,球體培養系統來源的MSC在抑制炎癥反應方面更有效。球體培養系統來源的MSCs治療組顯示血清中的中性粒細胞活性和促炎分子顯著降低 [70]。據程等人報道,注射到受損愈合傷口中的球狀MSC顯著增強了血管生成和愈合效率[66]。Bhang等人的研究證明,與對照組相比,球體培養系統來源的MSC移植組缺血區域觀察到更顯著的血管生成和更少的纖維化[74]。
然而,球體培養系統的一些缺點也限制了大規模生產MSC [75]。例如,球體培養技術會影響球體大小,而球體大小的可變性會影響MSCs的治療性能。因此,在臨床使用球體培養系統來源的MSC之前,有必要區分臨床需求,開發可重復的球體培養系統,并利用動物模型和臨床試驗確認其安全性和有效性。
腳手架
細胞外基質(ECM)被視為干細胞生存的“土壤”。生態位中的細胞通過整合素的跨膜蛋白機械錨定到ECM [76]。ECM的主要功能是為細胞生長和通過細胞-ECM相互作用,將機械傳感信號轉導至細胞以提供必要的支撐。它還為細胞提供必需的營養物質和生長因子 [61,77]。因此,有必要在viv中構建工程生態位來模擬原生ECM。生物材料的最新進展促進了MSCs人工ECM培養系統的發展,并且出現了各種天然和合成生物材料[61]。它們應滿足基質力學、可降解性和生物相容性等特性。現有的工程化ECM可以改善MSCs的生物學特性,例如增殖、歸巢、譜系分化和旁分泌[78,79,80,81]。
支架、薄膜、水凝膠和海綿形式的工程化ECM,已被開發并用于擴增MSC并增強其生物學特性 [82,83,84]。海藻酸鹽已廣泛用作組織工程中的生物相容性載體。Ewa-Choy等人的工作記錄了3D藻酸鹽水凝膠創造了一個微環境,促進ASC分化為軟骨樣細胞。ASC軟骨細胞分化的效率取決于藻酸鹽濃度[85]。值得注意的是,向ECM支架添加特異性誘導劑將進一步提高MSCs分化為所需細胞類型的效率。多項研究表明,羥基磷灰石(HAp)在骨重塑過程中與預先存在的礦物質相似,并具有優越的骨誘導活性 [86,87]。將BMSCs植入由鴨腳衍生膠原蛋白(DC)/ 含有或不含羥基磷灰石(HAp) 構建的多孔海綿中,以觀察它們的成骨分化。在骨傳導調節劑地塞米松的誘導下,DC/HAp海綿中的BMSCs表現出更高的增殖活性和成骨分化能力[88]。
接種在工程化ECM支架上的MSC的旁分泌特征和免疫調節作用也發生了顯著變化,接種在生物材料中的MSC顯示出PGE2和TSG-6等抗炎調節劑的上調。與2D培養體系中的 MSCs相比,海藻酸鹽包裹的MSCs可更有效地減弱TNF-α的分泌并增強PGE2的產生 [90]。此外,與單層培養相比,在藻酸鹽水凝膠中培養的ASC對植物血凝素刺激的外周血單核細胞的增殖有顯著抑制作用[91]。此外,海藻酸鹽水凝膠包封的MSC在體外促進巨噬細胞向抗炎M2表型的轉化。與對照組相比,它們在大鼠脊髓損傷 (SCI) 模型中發揮了類似的免疫調節作用,因為損傷部位的M2亞群百分比更高 [92,93]。
因此,與2D培養相比,基于支架的3D培養賦予MSCs更優異的生物活性和卓越的治療效果。隨著人們對生物材料的了解,其生物性能也在不斷擴大。僅使用生物材料,就可以制造出具有多種功能的仿生復合材料。例如,焦等人開發了一種具有明膠還原氧化石墨烯 (GOG) 和光交聯明膠水凝膠的雙相仿生原胼胝體,可提供缺氧微環境并介導BMSCs向成骨和血管生成的雙向分化,從而促進骨缺損的再生 [94]。因此,新一代基于支架的MSC培養技術不僅要同時從多個方面模擬干細胞生態位,提供盡可能多的干細胞生態位成分,還要釋放生物活性分子以靶向效應細胞,激活“自我修復”的機制。
o4
模擬病理微環境預激活
MSC進入人體后,將面臨疾病引起的病理微環境。在這種環境中存在多種破壞因素,可導致移植細胞的氧化應激和凋亡,并顯著損害MSCs固有的治療特性。研究人員致力于針對惡劣的病例環境微調細胞的特性,并使其更適用于特定的疾病。基于MSCs的可塑性和記憶能力,創傷微環境中的線索,如環境因素(缺氧)、化學因素(炎癥因子和細胞因子),將是MSCs體外操作的主要考慮因素 [95]。此外,MSC的制備工藝也在不斷探索求新,如與藥物、中藥和小分子化合物的結合也相繼出現 [28,96,97]。另一個值得關注的是,患者內源性MSCs的生物學特性會隨著疾病的發展而改變,表現出功能的喪失和衰退[98,99,100]。因此,提高間充質干細胞的生物學活性,提高其治療效果,特別是開發為患者定制的治療性間充質干細胞具有重要意義。
低氧預激活MSC
在前面的章節中已經提到,低氧是干細胞生態位的重要組成部分。實際上,在動物實驗和臨床研究中,間充質干細胞總是被輸送到缺血缺氧的損傷部位。因此,缺氧也是病理環境的重要組成部分 [34]。體外低氧培養可以誘導MSC對損傷微環境的記憶,從而提高治療效率,這在前幾節已有討論。然而,干細胞生態位和受損組織之間的氧濃度存在差異。此外,受損組織中的局部氧濃度在不同的修復期會有所不同。因此,干細胞預處理的氧濃度因目的而異。例如,MSCs的體外培養是指生理氧濃度,并在其體內應用之前用病理氧濃度進一步預處理。
用炎癥因子預激活MSC
炎癥因子和細胞因子的預激活被認為是模擬體內炎癥微環境的最常用手段,在調節干細胞的免疫調節功能方面發揮著重要作用[3]。TNF-α、INF-γ和IL-1β等促炎細胞因子經常在創傷性微環境中觀察到,并被廣泛研究用于MSCs的預激活[3]。
用TNF-α預激活MSC
越來越多的證據表明,用與疾病相關刺激對MSCs進行適當的啟動可以改善其生物學功能并發揮更好的治療作用[101]。TNF-α 在缺血和受傷組織中表達,通常用于模擬急性炎癥環境 [102]。用 TNF-α預激活牙齦組織衍生的MSCs (GMSCs) 可增強外泌體CD73的表達,這對于誘導抗炎M2巨噬細胞極化至關重要 [103]。來自TNF-α預活化的GMSC的外泌體表現出比對照更強的抗破骨細胞活性,從而減少結扎誘導牙周炎小鼠模型中的牙周骨吸收 [103]。此外,TNF-α預激活的MSC在H2O2誘導的氧化應激下顯著提高了增殖、遷移和存活率。并且它們通過大量分泌HGF、VEGF和其他細胞因子發揮比對照MSCs更好的內皮保護功能 [104]。此外,用TNF-α引發的MSC通過分泌促血管生成細胞因子(如IL-6 和IL-8)加速恢復了缺血性后肢和皮膚傷口的血運?[105,106]。
用干擾素 (IFN)-γ 預激活MSC
促炎細胞因子IFN-γ也是用于MSC預激活的代表性因子[107]。作為對IFN-γ的反應,MSCs具有獨特的免疫抑制特性,幾種抗炎因子的表達增加,例如HGF、TGF-β1、IDO、前列腺素和環氧合酶 2 (COX-2) [108,109,110]。由IFN-γ刺激的MSC分泌的前列腺素和 IDO是抑制NK活化的主要效應物[109]。此外,IL-2/15 激活的NK細胞對IFN-γ刺激的 MSCs的細胞毒性低于未刺激的MSCs,因為它們上調了抑制性MHC I類分子,而IFN-γ引發的MSCs抑制PBMCs增殖的能力比未刺激的MSCs更強[111],并伴隨著PD-L1的上調和COX-2衍生的PGE2分泌的增加[112]。在CCl4誘導的肝硬化 [113]、閉塞性細支氣管炎[114] 和腎纖維化[115]模型中,證明MSCs在IFN-γ預激活后的治療潛力得到顯著改善。這一證據表明,間充質干細胞可以被炎癥信號激活,并足以增強其免疫調節特性和治療效果[110]。
用IL-1β預激活MSC
IL-1β也是由單核細胞和巨噬細胞產生的炎癥組織中普遍存在的炎癥細胞因子 [116]。研究表明,IL-1β預激活增加了MSCs中許多粘附分子的表達,例如整合素 LFA-1,從而通過與ICAM-1的相互作用促進對HUVECs的粘附,從而促進MSC跨內皮細胞和歸巢 [117]。此外,Nie等人的研究發現,IL-1β預激活的MSCs表現出升高的CXCR4表達并增加了它們向SDF-1的遷移,導致在急性肝功能衰竭中比初始MSCs具有更好的治療性能 [118]。此外,來自IL-1β預激活的MSCs的外泌體可以誘導巨噬細胞極化為M2表型,并且比幼稚MSCs產生的外泌體更有效地減輕膿毒癥小鼠模型中的癥狀 [119]。
用其他促炎細胞因子和TLR配體預激活MSC
此外,最近報道了一些其他炎癥因子如IL-17A和IL-25作為替代的預激活手段并獲得了有希望的結果。IL-17A主要由CD4+T輔助細胞 17 產生,并在發展自身免疫和炎癥性疾病中起調節作用 [120]。一項比較研究的結果表明,用 IL-17A 刺激的 MSCs 表現出比未處理的 MSCs 更好的免疫抑制特性 [121]。與 IFN-γ、TNF-α 或 IL-1β 處理的MSCs 相比,IL-17A 處理的MSCs對有絲分裂原活化的CD3 + T細胞的抑制程度最高。IL-25 是細胞因子 IL-17 家族的成員,最近被用于增強MSC調節的免疫反應[122,123]。與未引發的 MSC相比,注入IL-25引發的MSC顯著減少了IL-17陽性細胞并增加了FoxP3陽性細胞,從而減輕了 DSS 誘導的結腸炎大鼠模型中的腸道炎癥 [122]。
脂多糖 (LPS) 是革蘭氏陰性菌外膜的一種成分,在各種疾病中升高。最近的研究表明,LPS 是調節多種細胞類型細胞凋亡的重要介質 [124]。已經證明 LPS 對 MSCs 的生物學效應與使用的LPS濃度密切相關。高劑量LPS處理的MSCs誘導其凋亡,低劑量LPS處理的MSCs增強了其抵抗氧化應激和抑制細胞凋亡的能力,可能取決于細胞FADD樣IL-1β轉化酶抑制蛋白的上調. 此外,通過MyD88依賴性激活stat3 [124,125,126],移植低劑量LPS預活化的MSCs在 I/R損傷模型中顯著改善了MSC介導的心臟保護作用。
用促炎細胞因子組合預激活 MSC
間充質干細胞經多種促炎因子預激活后,其生物學活性發生變化,因此科學家們設想是否可以通過聯合不同的炎癥細胞因子對間充質干細胞進行預處理,以彌補效率低下,最大限度地發揮治療作用。因此,炎性細胞因子的預激活不僅可以提高間充質干細胞抵抗氧化應激的能力,而且在很大程度上增強間充質干細胞的免疫抑制特性,增強其治療效果。盡管如此,仍有一些問題需要回答。需要深入研究并進一步探索和確定單獨或組合炎性細胞因子預激活的最佳濃度和作用持續時間。或者,還應關注炎癥預激活的可能副作用,例如I類和II類HLA分子的不良上調。
用生長因子或再生細胞因子預激活 MSC
最近出現了用生長因子或再生細胞因子激活MSC,并已被證明是一種有吸引力的方法。用混合生長因子預激活 MSCs 顯示出增強其生物學功能的協同作用。此外,與移植未經處理的MSCs相比,這些預激活的MSCs的移植可減少梗塞面積并改善心臟功能 [136]。然而,為預激活的 MSCs 選擇最佳細胞因子并賦予所需的生物學功能是重要的一步。是選擇在各種疾病中共同變化的細胞因子,還是使用疾病特異性細胞因子作為預激活條件,是一個需要深入研究的問題。或者,細胞因子預激活的方法不夠經濟,因為大規模生產預激活的MSC需要大量的細胞因子。
用生物活性化合物預激活MSC
生物活性化合物是一種很有前途的預活化方法,可增強MSCs的生物學特性 [28,96,137]。目前用于刺激間充質干細胞的生物活性化合物按其來源可分為天然(如從中藥中提取)和合成化合物;在生物活性化合物的篩選原理上,要么對MSCs有生物調節作用,要么對目標疾病有治療作用;鑒于其對間充質干細胞的生物學機制,生物活性化合物可分為以下幾類:促進存活和遷移,增強分泌活性,恢復和修復失能的間充質干細胞。
o5
促進間充質干細胞存活和遷移的生物活性化合物
實際上,發揮 MSCs 固有的治療特性需要移植的細胞在嚴酷和受損的環境中存活并發揮作用[28]。許多研究都集中在通過使用生物活性化合物來修飾 MSCs 以增強其抗凋亡和遷移能力。
曲美他嗪(TMZ)可降低缺血引起的組織損傷,常用于治療心絞痛。它通過增加促生存因子如 HIF-1α、Akt、survivin 和 Bcl-2 的表達來保護MSCs免受過氧化氫 (H2O2) 誘導的氧化應激。他達拉非屬于長效PDE5抑制劑組,已用于治療心力衰竭 [138]。它通過上調miR-21以抑制Fas [138,139] 來改善MSCs體外的增殖和存活。此外,阿托伐他汀對內皮功能產生有益的影響[140],促進了MSCs的存活,并通過eNOS/NO和SDF-1/CXCR4 通路促進MSCs 在梗塞心臟中的治療作用 [140]。
維生素E是一種眾所周知的抗氧化劑,因為它具有清除自由基的活性 [141]。維生素E預激活的MSC對H2O2誘導的氧化應激以及增殖標志物(增殖細胞核抗原和Ki67)和促再生標志物(TGF-β和VEGF)的上調具有抗性。此外,移植含有維生素E的MSC 可修復大鼠骨關節炎模型中受損的軟骨 [142]。
近年來,已經研究了中藥或其提取物對 MSCs 的有益作用 [143 ]。丹參 (SM) 是一種廣為人知的草藥,常見于許多中醫處方中,用于治療各種疾病,包括心血管疾病、阿爾茨海默氏癥和缺血性中風[144,145,146]。SM有效地增強了MSCs在缺氧條件下的活力并減少了細胞損傷。在大鼠大腦中動脈閉塞模型中,SM修飾的MSCs的輸注顯示了梗塞區域的恢復和積極的行為變化 [147]。姜黃素是姜黃 (Curcuma longa) 的活性成分,具有抗氧化和抗炎等多效作用 [148,149]。它通過調節PTEN/Akt/p53 通路和血紅素加氧酶1的表達對ASC中的氧化應激誘導的損傷發揮細胞保護作用[150,151]。先前的姜黃素治療顯著增加了 MSCs中VEGF的分泌,并且這些預激活的MSCs在缺血性心肌中比初始ASCs促進了更多的新血管形成和功能恢復 [152]。
此外,姜黃素預活化的MSCs改善了其在酸性燒傷傷口中的治療潛力,表現出改善微循環、明顯的肉芽形成和加速傷口閉合的潛力?[153]。還有許多其他中藥提取物,如迷迭香酸 (RA) 和 gigantol,它們通過調節caspase-3、caspase-9和Bax/Bcl-2的表達來保護MSCs免受H2O2誘導的細胞凋亡。PI3K/Akt和ERK1/2信號通路 [154,155,156]。這些結果表明它們可以作為細胞保護劑用于MSC移植。
用于增強MSCs的免疫調節、旁分泌和治療潛力的生物活性化合物
事實上,間充質干細胞的免疫調節和旁分泌特性與其治療效果密切相關。最近的數據表明,藥理刺激可以增強MSCs的旁分泌和免疫調節潛力。
鐵螯合劑去鐵胺 (DFX) 是一種具有抗氧化特性的低氧模擬劑。用于預激活MSCs的DFX導致抗炎(IL-4、IL-5和COX2)、促血管生成因子(VEGFα 和血管生成素-1)以及神經保護因子(神經生長因子、間充質干細胞中的膠質細胞衍生的神經營養因子和神經營養因子-3) [157]。DFX預激活的MSC的分泌組可以有效地將LPS誘導的巨噬細胞 DH82 重編程為M2表型[158]。它還顯示出背根神經節 (DRG) 神經元在高劑量葡萄糖誘導的損傷下的神經保護潛力 [157]。Treprostinil 是一種前列環素類似物,用于刺激 MSCs 并通過增加 VEGF-A 的產生來發揮促血管生成作用 [159 ]。在全反式維甲酸 (ATRA) 中發現了促生存、血管生成和促遷移相關基因的表達,包括 COX-2、HIF-1、CXCR4、CCR2、VEGF、Ang-2 和 Ang-4經處理的MSCs [160]。此外,與注射未經處理的MSCs 的傷口相比,注射ATRA處理的MSCs的傷口顯示出顯著更高水平的血管化、膠原沉積和再上皮化,促使傷口加速閉合 [160]。白藜蘆醇 (RSV) 是一種植物多酚類化合物,可保護MSCs免受炎癥和氧化損傷 [161,162],增強其旁分泌功能,防止其衰老等 [163,164]。例如,RSV預激活增強了MSCs中 PDGF-DD 的分泌,進一步激活了腎小管細胞中的ERK信號通路,促進了內皮細胞的血管生成,并優選修復了順鉑誘導的腎損傷 [164]。補陽還五湯(BHD)是中醫補氣活血的名方,用于治療中樞神經疾病[165]。與未經處理的 BMSCs 相比,BHD預激活的大鼠 BMSCs 衍生的外泌體含有更多的血管生成miRNA,并且在雙側頸動脈結扎后大鼠腦中的血管生成增加 [166]。
高通量技術的發展為我們篩選靶向MSC中特定基因的生物活性化合物,從而調控MSC的表達譜帶來了新的視角。例如,漢防己甲素被選擇來特異性上調 PGE 2 的表達。MSCs通過 NF -κ B / COX – 2 信號通路預激活的四氫萘MSCs在與小鼠巨噬細胞( RAW 264.7)共培養后, TNF -α分泌顯著減少,小鼠炎癥耳朵中腫瘤壞死因子-α水平減弱[167].
用于逆轉和修復功能殘缺的MSCs的生物活性化合物
圍產期組織間充質干細胞在體外大規模擴增后會發生復制性衰老,老年供體間充質干細胞普遍呈現早衰表型,患者的間充質干細胞表現出MSC生物學功能下降。這些MSC通常在增殖、遷移、分化和免疫調節方面表現出細胞功能下調,同時治療能力受損 [98,99,100]。因此,逆轉這些MSCs的功能,即MSCs的再生,對于基于MSC的治療至關重要。
最近的研究表明,細胞功能下降或過早衰老可以得到挽救 [168]。一般來說,拯救 MSCs 的方法可以廣義地定義為降低細胞內氧化應激水平,通過調整表觀遺傳修飾對 MSCs 進行重編程,以及使用抗衰老藥物。
抑制過度氧化應激
數據表明,作為細胞代謝產生的天然副產物,ROS 在 MSCs 中維持在低水平,對 MSCs 的增殖和分化至關重要 [169,170]。在實際老化或病理條件下,高水平的ROS誘導會導致嚴重的細胞毒性和細胞損傷 [168]。幾項研究報道,通過調節細胞內 ROS 聚集和氧化代謝可以逆轉發生故障的MSCs [168]。抗氧化劑已成為自然選擇,并且有幾種抗氧化劑用于 MSCs的抗衰老研究。N-乙酰半胱氨酸 (NAC) 是一種ROS清除劑,可顯著減弱MSCs中 Wnt/β-catenin 信號傳導過度激活導致的ROS積累,從而減輕ROS誘導的DNA損傷并下調衰老相關標志物p16 (INK4A)、p53 的表達和p21 [171]。另一種自由基清除劑依達拉奉通過降低 ROS 水平和 β-gal 陽性細胞來挽救老年 AT-MSCs 的功能。此外,它還可以保護BMSCs免受缺氧和UC-MSCs中抗氧化酶上調引起的細胞內ROS積累 [172,173]。更重要的是,用依達拉奉預激活恢復了老年 AT-MSCs 的體內治療功能,因為缺血皮瓣小鼠模型中的壞死面積減少了 [174]。
總體而言,ROS 可以作為細胞內信使并幫助執行重要的生物學功能,因此必須通過調節抗氧化劑的量來控制 ROS 的最佳濃度。畢竟高劑量的抗氧化劑會導致DNA損傷和過早衰老[175]。
用于修飾表觀遺傳失調的生物活性化合物
表觀遺傳調控是一種重要的編程機制,它通過改變基因表達而不是DNA序列來改變細胞表型。它具有遺傳性和可逆性的特點,包括 DNA 甲基化和組蛋白修飾,對MSC命運有深遠的影響 [176,177]。常規培養擴增后在 MSCs 中發現的表觀遺傳失調似乎與整體組蛋白乙酰化水平的變化無關,但涉及某些基因啟動子的組蛋白乙酰化水平,如TERT、Soc2、Oct4、Runx2 和 ALP,最終導致到細胞衰老 [178]。此外,通過使用BeadChip微陣列,與來自年輕的 MSCs 相比,據報道,來自老年的MSCs的DNA甲基化普遍降低 [179]。鑒于表觀遺傳修飾的可逆性,探索表觀遺傳靶向治療將老干細胞重編程為年輕的功能性干細胞是一種潛在的策略。
使用目前的技術,對衰老干細胞進行重編程的方法主要有兩種,即完全重編程和部分重編程。前者是指重置表觀遺傳時鐘以最終獲得誘導多能干細胞(iPSCs),但它們抑制T細胞增殖的免疫調節功能不完整 [180,181,182]。因此,需要徹底評估iPSCs衍生的MSCs的治療效率,尤其是免疫調節功能。此外,由于效率低、iPSC數量有限、成本高,完全重編程在實驗室還只是一種手段,離床邊還有相當大的距離。部分重編程涉及不完全去分化,被認為是表觀遺傳返老還童,可以通過使用生物活性化合物調節DNA甲基化和組蛋白修飾來實現 [183]。
組蛋白乙酰化增加、DNA 甲基化和羥甲基化減少以及基因組中 H3K27me3 的明顯變化在衰老細胞中普遍存在。然而,通過改變表觀遺傳修飾來逆轉干細胞衰老仍處于起步階段。有必要繪制出衰老過程中MSCs 的詳細表觀遺傳變化,特別是與MSC生物學特性改變相關的表觀遺傳特征。此外,不同來源的MSC老化表觀遺傳變化的普遍性或獨特性需要得到證實。
抗衰老藥物的使用
Senotherapeutics 是指應用抗衰老藥物選擇性地消耗衰老細胞或延緩衰老的發生,從而使組織恢復活力并減少與年齡相關的病變的發生 [192]。據報道,有幾種化合物具有潛在的抗衰老作用,例如納維他 (ABT-263)、槲皮素、達那唑、煙酰胺核苷、達沙替尼和二甲雙胍 [192,193]。用抗衰老藥物達沙替尼治療,既能選擇性地促進衰老MSCs的凋亡,又能顯著改善 MSCs的生物學活性[194]。
可以想象,不同藥物的組合可能會達到更明顯的抗衰老作用。如Zhou等人的研究所示,達沙替尼和槲皮素的抗衰老混合物在體外或體內顱骨缺損模型中,改善了老年小鼠來源的骨髓間充質干細胞的成骨潛力[195]。值得注意的是,一些研究結果表明,達沙替尼和槲皮素的混合物可以減輕衰老細胞的負擔,并改善脂肪、骨骼、主動脈和大腦等重要組織的功能 [195,196]。然而,需要更多的證據來支持senolytic雞尾酒療法在挽救MSCs功能方面的有效性。
一般來說,這些發現表明生物活性化合物具有改善功能確實的MSC和修復衰老MSC的特性,開發更有效的生物活性化合物來拯救MSCs也是科學家們需要繼續努力的方向。此外,生物活性化合物的使用方式也值得仔細考慮。正如幾項研究所證明的,使用生物活性化合物作為 MSC 移植的伴隨藥物,也比單獨的干細胞移植具有更好的治療效果[24,95,197]。盡管如此,慢性病患者還伴隨著多種危險因素,例如年齡、糖尿病和心血管疾病。因此,應考慮干細胞移植伴隨藥物的可能副作用和安全性。MSCs的衰老是一個高度復雜的過程,深入了解衰老的潛在機制,將有助于我們找到更有效的方法來使衰老的MSCs恢復活力。
用疾病相關效應細胞或患者血清預激活 MSC
“個體化MSCs治療”是指通過體外預激活獲得的MSCs具有定制化的功能,可以特異性針對患者的疾病,從而達到更好的治療效果。因此,增強MSCs的治療效果,建議直接使用效應細胞或其釋放的活性物質作為預激活條件,而不是使用典型的促炎因子、細胞因子或生物活性化合物進行教育。
肥大細胞 (MC) 在速發型超敏反應和過敏反應中起核心作用,也是特應性皮炎 (AD) 發病機制中的主要效應細胞。活化的 MC 釋放含有大量生物活性物質的顆粒,例如促炎細胞因子、蛋白質介質、脂質介質和生長因子,可引發過敏反應 [199]。多項研究表明,MSC 在共培養系統中抑制MC活化和脫粒并誘導MC凋亡 [200,201,202]。然后,具有MC顆粒的預激活 MSCs 可能是增強 MSC 靶向治療 AD 的一種有前途的策略。Lee等人的研究表明,MC顆粒引發的 UC-MSCs 比未引發的細胞表現出更強的免疫抑制性,這是通過上調 COX-2/PGE2 信號通路來中斷MCs的增殖和脫粒來介導的[201]。此外,在 dermatophagoides farina 誘導的 AD 模型中,與幼稚細胞相比,皮下輸注MC顆粒培養的UC-MSC顯示出更顯著的MC數量減少并減輕了皮膚中淋巴細胞的浸潤 [201]。因此,用效應細胞或其衍生活性物質預激活的間充質干細胞,可以準確靶向疾病發展過程中的主要致病因素,并在體內快速反應和響應,從而達到更有效的治療效果。
血液循環中炎癥因子、趨化因子、生長因子、細胞因子和微泡的改變已在多種疾病中發現,包括阿爾茨海默病、腎病和心臟病[203,204,205]。用患者來源的血清預激活擴增的MSCs 可以使MSCs對宿主微環境做出積極反應 [203]。Tang等人的研究表明,與注射對照血清預激活MSCs的大鼠相比,注射中風血清預激活MSCs的大鼠表現出行為顯著改善,炎癥細胞因子減弱,腦損傷和凋亡細胞減少,和增加的腦I/R損傷模型中的營養生長因子[206]。此外,在葡聚糖硫酸鈉誘導的結腸炎大鼠模型中也顯示出用疾病衍生血清增強 MSCs 的治療效果。MSCs用結腸炎大鼠或正常大鼠血清預活化后,收集兩種預活化細胞的條件培養基用于治療結腸炎。前者顯示出比后者更加有效地阻止疾病進展,更好地改善臨床特征,并且在結腸炎大鼠中的組織學損傷評分低得多[207]。
因此,疾病特異性預激活可能是實現“MSC定制臨床治療”的有希望的手段。而準確捕捉疾病特異性是該策略的必要前提,需要對疾病的生物學機制進行全面深入的探索和分析。
看法
除了通過重建生理和病理微環境來預激活MSCs外,還有其他方法可以預激活MSCs,包括光刺激、磁電刺激和熱休克(HSP)等。低水平激光治療(LLLT)有利于調節多種細胞的生物學功能 [208,209]。例如,與未受刺激的ASC相比,低強度激光刺激的ASC的生物活性增強,表現為提高存活率、增加分泌和加速再生愈合 [210,211]。此外,最近已證明脈沖電磁場通過調節 Akt/Ras 信號通路和上調 Bad 和 Bcl-xL 等存活蛋白對 BMSCs 發揮保護作用 [21]。此外,研究表明,HSP可以誘導細胞保護蛋白并增加抵抗惡劣外部環境的能力 [213,214]。HSP預激活增強了MSCs自噬并增加了它們對H2O2誘導的細胞凋亡的抵抗力。此外,與對照MSCs相比,HSP-MSCs在肝I/R 損傷模型中移植后顯示出增強的歸巢特性和存活率 [215]。此外,卵巢內注射 HSP-MSC 可挽救受損的卵巢結構并改善內分泌功能 [216]。
一般而言,現有預激活方法的目的是充分利用MSCs的功能可塑性,提前為MSCs賦予所需的特性,以便MSCs再次遇到類似環境時,可迅速響應,啟動細胞保護機制。
盡管如此,預激活 MSCs 的應用面臨著一些挑戰。
一是選擇合理有效的MSC預激活方法。迄今為止,已經提出了許多補充方法來提高MSCs的治療效果,尋找最佳的預激活方式是我們未來研究的重要追求。每種預處理都有望改善MSCs 在臨床應用上的效果,例如,將 MSCs 接種到與 bFGF 或 TGF-β1 交聯的膠原微載體的可注射凝膠中,以促進其定植和向成軟骨分化[217]。此外,基于效應細胞或基于患者血清的 MSCs 預激活可能相對更有針對性地用于疾病治療,這需要大量的比較研究來進一步確定這一推論。
二是間充質干細胞的異質性,主要表現在兩個方面:一是不同來源間充質干細胞的生物學特性存在差異,這意味著作為一種新型細胞藥物,間充質干細胞來自不同組織對同一疾病的治療效果不一致。
此外,當面對相同的預激活條件時,來自不同組織的 MSC 的反應機制會有所不同。例如,刺激/未刺激的 UC-MSCs 與植物血凝素激活的淋巴細胞共培養導致 UC-MSCs 中負共刺激分子 CTLA4 的早期激活,而在共培養的BMSCs中改變了IL-12的表達[219]。這些差異可能部分由來自不同組織的MSC之間的功能異質性決定。
三是開發標準化的生產平臺,用于評估預激活MSCs的安全性和治療特性(圖3)。大量研究證據證實,間充質干細胞主要依靠旁分泌發揮生物學作用。MSCs分泌產物主要包含可溶性蛋白和細胞外囊泡,前者包括細胞因子、趨化因子、生長因子,后者可分為外泌體和微泡[220,221](圖1)。因此,準確選擇成分作為受過“教育”的MSC的預激活和治療效果的指標是一個棘手的問題。然而,越來越多的證據表明,移植的MSCs的體內凋亡與多種動物模型中的干細胞治療效果密切相關,例如GvHD [222]、敗血癥和急性肺損傷 [223,224]。Pang等人的研究結果進一步說明通過消融BAK/BAX來阻止MSC凋亡會降低它們在OVA誘導的哮喘模型中的免疫調節能力,這表明MSCs的體內生物學機制遠比我們想象的要復雜得多 [225]。因此,抑制和抵抗干細胞凋亡作為MSC預激活的常用評價指標值得商榷。
?
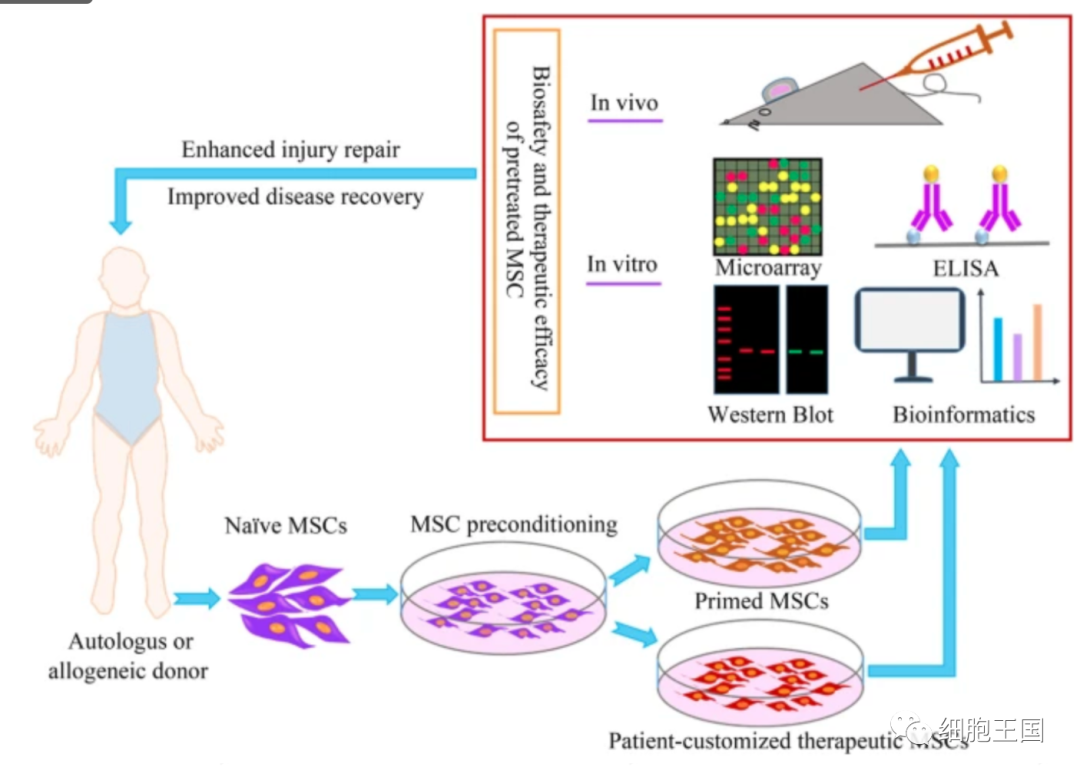
預處理的 MSCs 取得了良好的治療效果,增強了損傷修復和疾病治療的能力。
最后,MSC體內治療的本質是遞送促修復調節因子和細胞外囊泡等。因此,研究人員開始使用MSC衍生的可溶性蛋白和細胞外囊泡替代干細胞進行治療[226,227],其釋放受到精確調節,其成分隨預激活條件而變化 [228]。該方法可以避免MSCs在體內增殖、分化和分泌不可預測的旁分泌因子的風險。
最近的結果表明,MSCs通過密度梯度離心被去核成“cargocytes”,形成一種仿生遞送方法。Cargocytes 保留旁分泌能力,且不增殖,或可永久移植到宿主中。Cargocytes 的應用不僅避免了與直接使用MSCs相關的不良事件,例如肺或腦栓塞,而且還改善了生物分布并增強了對體內靶組織的歸巢[229]。因此,來自MSCs的衍生物可能是一種可控的治療載體,將成為治療疾病的潛在物質。
結論
總之,盡管預激活的MSCs仍然存在問題,但由于其巨大的再生潛力,它們仍然對治療各種難治性疾病具有相當大的前景。迄今為止,越來越多的共識是,在各種病理條件下,預激活的MSCs確實比初始MSCs表現出更好的治療效果。
未來,我們應有效利用“預激活”工具,一方面最大限度地發揮MSCs的治療潛力,另一方面對其進行修飾使其適合靶向疾病,開啟MSCs臨床應用的新篇章。
—?THE END —
編輯:小果果,轉載請注明出處:http://www.448371.com/zixun/hydt/13793.html
免責聲明:本站所轉載文章來源于其他平臺,主要目的在于分享行業相關知識,傳遞當前最新資訊。圖片、文章版權均屬于原作者所有,如有侵權,請及時告知,我們會在24小時內刪除相關信息。
說明:本站所發布的案例均摘錄于文獻,僅用于科普干細胞與再生醫學相關知識,不作為醫療建議。
 微信掃一掃
微信掃一掃  支付寶掃一掃
支付寶掃一掃 
請問作者文中的參考文獻清單在哪兒可以下載???
@303775206:網上轉載文章,原文沒有參考文獻出處