

Treg細胞是CD4+T細胞的一個亞群,通過預防自身免疫、控制過度炎癥和維持免疫耐受,對免疫穩態至關重要。Treg細胞的免疫調節特性在癌癥中會促進腫瘤的進展。對TIL亞群的分析表明,免疫細胞浸潤的組成與患者預后之間存在明確的關聯。過表達細胞毒性T細胞的TIL通常與良好的預后相關,而TIL中Treg細胞的頻率增加與不良預后相關。
Treg細胞的分化和功能依賴于譜系特異性轉錄因子Foxp3的表達。FOXP3基因功能喪失改變的人類患有免疫失調、多內分泌疾病、腸病、X連鎖(IPEX)綜合征,表現為廣泛的自身免疫和炎癥損傷。Treg細胞在胸腺中由一組前體細胞發育而來,這是一個需要T細胞受體(TCR)和IL-2激活的兩步過程。
Treg細胞可以通過各種不同的機制抑制免疫反應。CD25是IL-2受體的一種高親和力亞單位,在Treg細胞上組成性高表達,但在效應T細胞上也上調。Treg細胞上高水平表達的CD25競爭性地結合IL-2,從而剝奪效應T細胞的這種專性生長因子。Treg細胞還表達一系列抑制分子,如CTLA-4,與Treg介導的抑制有關。此外,Treg細胞還可以分泌抑制性細胞因子,如IL-10和TGF-β。
基于這些有效的免疫抑制機制,Treg細胞通常被認為是最有效的抗腫瘤免疫抑制劑。因此,以Treg細胞為靶點的癌癥治療策略也是目前的研究熱點。
Tregs可分為兩類:天然調節性T細胞(nTregs)和誘導調節性T細胞(iTregs)。兩種類型的Tregs都普遍表達 Foxp3。nTregs在胸腺中自然發育,其抑制作用是通過細胞間接觸實現的,它們的主要功能是維持正常的免疫耐受和控制炎癥反應。
iTregs來源于腫瘤微環境信號誘導的外周原始T細胞,包括腫瘤抗原、細胞因子(如TGF-β)和其他可溶性分子。iTregs通過多種促進腫瘤進展的機制抑制效應T細胞(Teff)、NK細 胞和DC的抗腫瘤免疫作用。
Tregs主要有以下五種功能機制:①Tregs分泌抑制性細胞因子,包括 IL-10、TGF-β 和 IL-35。② Tregs 通過顆粒酶和穿孔素殺死效應細胞。③ Tregs 影響效應細胞的功能:Treg 與效應T細胞競爭消耗 IL-2,從而抑制效應T細胞的生長;Tregs通過產生胞外酶CD39和CD73 促進TME中腺苷的產生,誘效應細胞導的抑制和抗增殖作用;Tregs通過縫隙連接將大量 cAMP 轉移到效應T細胞,干擾其代謝。④Tregs通過刺激性和抑制性受體(CTLA-4 或 LAG3)誘導DC耐受,后者通過IDO進一步抑制T細胞的能力。⑤MDSCs和Tregs 產生的因子形成正反饋環,促進增殖,增強抑制環境。? ? ? ? ?
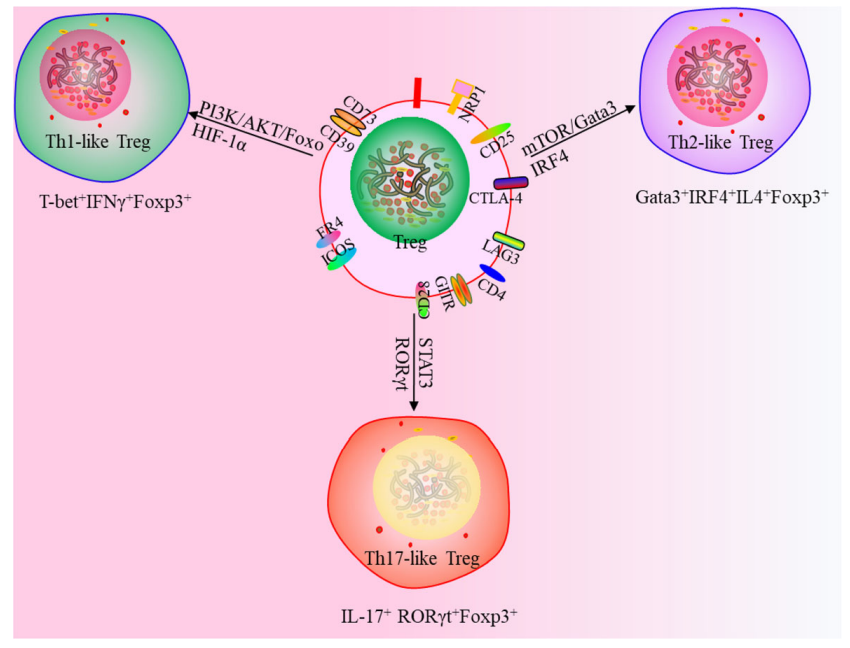

此外,一項研究使用一種新的策略來定義Tregs, Th1樣Tregs(T- 234bet+IFNγ+Foxp3+),Th2 樣Tregs(Gata3+IRF4+IL4+Foxp3+)和Th17樣Tregs(IL-17+RORγt+Foxp3+),這為靶向 Tregs 治療提供了新思路。
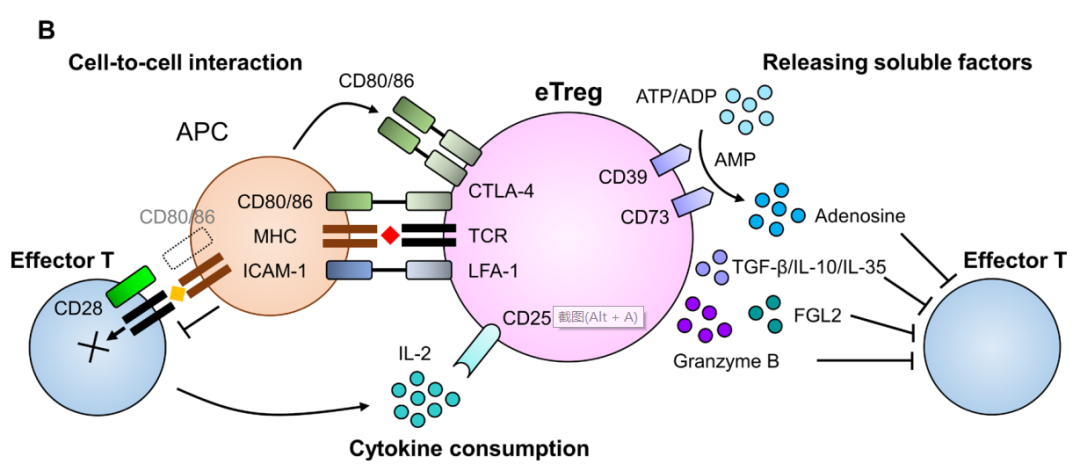
Treg通過多種機制阻礙抗腫瘤免疫反應。在Tregs中,共抑制受體細胞毒性T淋巴細胞抗原4(CTLA-4)通過與APCs中的CD80/B7-1和CD86/B7-2結合干擾共刺激信號,從而抑制效應T細胞中的共刺激受體CD28。? ? ? ? ? ?
 ? ? ? ? ? ? ??
? ? ? ? ? ? ??
IL-2是Treg和效應T細胞生存所必需的細胞因子。與效應T細胞相比,Treg主要通過由α(CD25)、β(CD122)和γ(CD132)亞單位組成的更高親和力受體結合IL-2。此外,Tregs比效應T細胞具有更高的CD25表達,而CD25是IL-2的一種高親和力受體。這使得Treg在利用TME中有限的IL-2方面具有競爭優勢。因此,在TME中,Treg的累積量大于效應T細胞。同時,Treg還分泌免疫抑制分子(包括免疫抑制細胞因子)抑制抗腫瘤免疫。Treg產生TGF-β、IL-10和IL-35導致免疫抑制。
表達在Tregs細胞表面的CD39和CD73作為外核苷酸酶,分別將ATP或ADP水解為AMP和AMP水解為腺苷。CD39和CD73產生的腺苷會抑制效應T細胞。當Treg在TME中發生凋亡時,凋亡的Treg通過外核苷酸酶釋放大量腺苷,從而產生更強的抗腫瘤免疫抑制。
趨化因子和細胞因子依賴性浸潤
Treg具有多種趨化因子受體。趨化因子梯度,如CCR4-CCL17/22,CCR5-CCL5, CCR8-CCL1,和CCR10-CCL28,可參與將Treg招募到TME中。? ? ? ? ? ? ? ?

在炎癥腫瘤中,炎癥細胞產生Treg招募趨化因子,如CCL22。CCR8+Treg被CCR8配體(如CCL1和CCL18)招募到TME等炎癥部位,CCL1不僅將CCR8+Treg招募到腫瘤中,還誘導對Treg抑制至關重要的FOXP3、CD39和IL-10的STAT3依賴性上調,從而增強Treg的免疫抑制活性。在一系列細胞因子和趨化因子受體中,與正常組織駐留的Tregs相比,CCR8僅在人類乳腺癌的腫瘤浸潤性Tregs中顯著上調,表明CCR8是TME中Tregs的一個有希望的治療靶點,而不會引起系統性自身免疫。
基于TME代謝適應的生存優勢
效應T細胞和Treg在正常和炎癥條件下使用不同的代謝系統,Treg表現出更強的生存優勢。
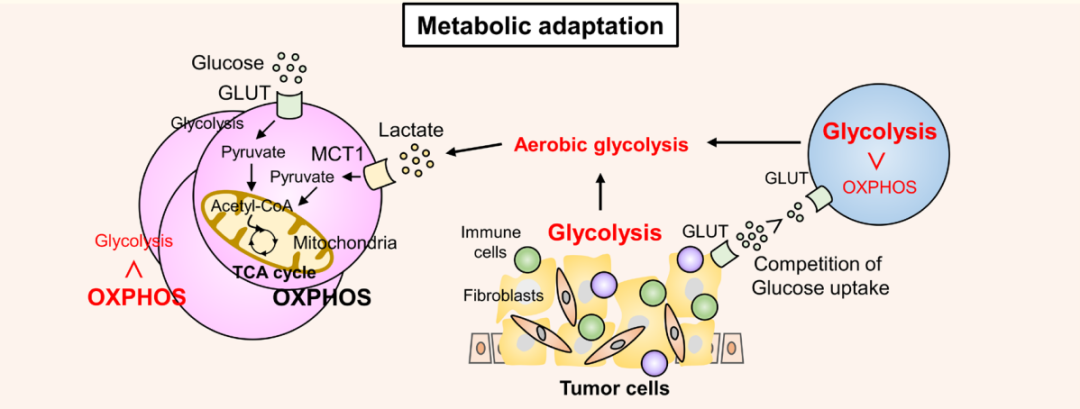
TCR刺激通過磷酸肌醇3-激酶(PI3K)-雷帕霉素(mTOR)信號途徑激發特定的代謝程序,導致氨基酸和葡萄糖的吸收增加,從而增強有氧糖酵解。活化的效應T細胞將其代謝程序從氧化磷酸化(OXPHOS)轉變為有氧糖酵解,這是其生存和功能所必需的,從而導致效應T細胞和TME中的腫瘤細胞之間的葡萄糖競爭。腫瘤細胞中的代謝重編程將TME轉變為營養受限,富含乳酸和低氧環境,不利于效應T細胞的存活和功能。
然而,在TME中,Treg可以在如此惡劣的條件下存活并保持其免疫抑制功能。FOXP3通過抑制糖酵解和促進氧磷和煙酰胺腺嘌呤二核苷酸(NAD+)氧化,因此,Treg可以利用乳酸作為能量來源。當腫瘤浸潤性非Treg將丙酮酸轉化為乳酸以產生NAD+以維持糖酵解時,TME中的Treg將丙酮酸轉化為線粒體中的乙酰輔酶a以觸發三羧酸循環,這為Treg提供了比其他T細胞更大的存活益處。
此外,即使在低葡萄糖條件下,Treg還可以利用脂肪酸實現其增殖和免疫抑制功能。研究發現,胃癌中RHOA Y42突變通過上調脂肪酸合成酶(FASN)產生大量脂肪酸,導致TME中更多的Treg和更少的腫瘤浸潤性CD8+T細胞。
非炎癥腫瘤的浸潤機制
雖然炎癥腫瘤通常含有Treg,但有時在非炎癥腫瘤中也檢測到大量Treg,這表明炎癥相關浸潤以外的機制參與了Treg向TME的募集。
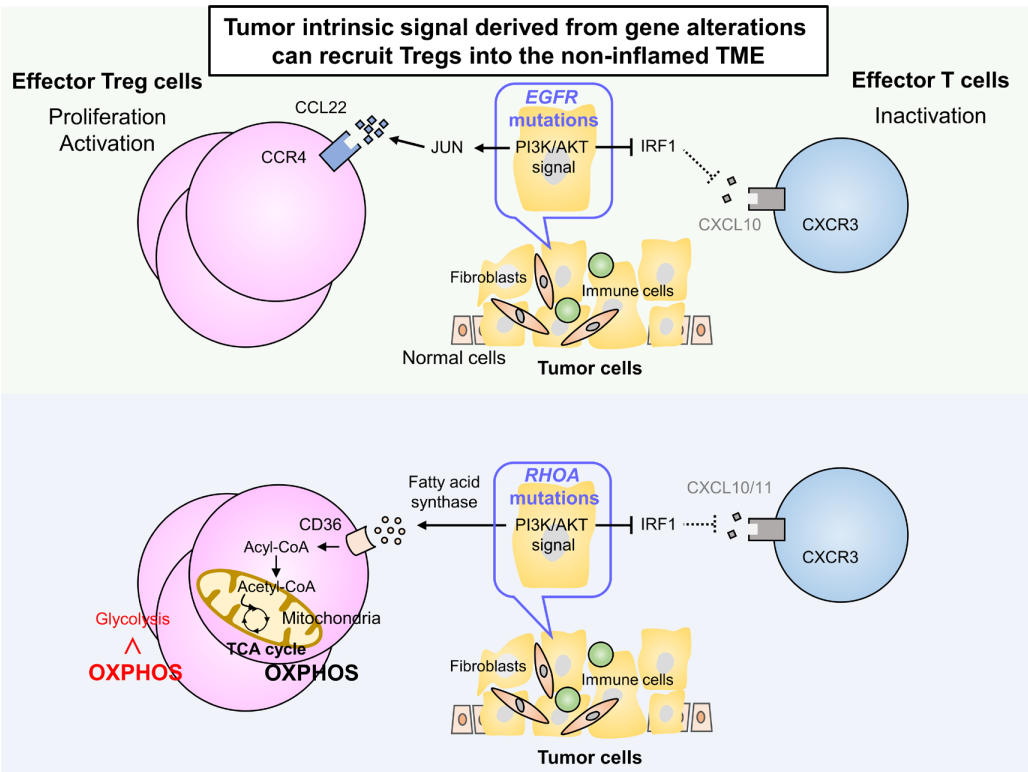
研究發現,某些基因改變可以通過調節下游信號通路來修飾腫瘤細胞產生趨化因子。在肺腺癌中發現的EGFR突變通常與非炎癥性腫瘤相關,在這些腫瘤中檢測到大量的Treg,而不存在炎癥細胞。
EGFR突變通過抑制IFN調節因子1(IRF1)減少CXCL10的產生,這對CXCR3依賴的CD8+T細胞向TME的募集產生負面影響。此外,CCL22的產生通過EGFR下游信號中的JUN誘導而增加,導致CCR4依賴性Treg浸潤到TME。
在具有RHOA Y42突變的胃癌中也觀察到類似的表型,RHOA Y42突變是一種功能缺失突變,通過抑制IRF1降低CXCL10/11水平。此外,粘著斑激酶的過度激活通過調節趨化因子的產生與Treg浸潤和CD8+T細胞排斥相關。
治療相關的Treg積累
放射治療和CTLA-4阻斷抗體等治療可能促進腫瘤中Treg的積累,這被認為是一種適應性免疫抵抗機制。由于對輻射的敏感性低于其他淋巴細胞,因此即使在放射治療后,Treg仍在TME中持續存在,可能導致某些類型癌癥發生免疫抑制性TME。在臨床前模型中,去除Treg可提高腫瘤對輻射的敏感性并抑制轉移。
在小鼠模型中,抗CTLA-4抗體通過抗體依賴性細胞毒性(ADCC)有效地去除Treg,而在患者中的作用機制仍存在爭議。免疫組化分析顯示,根據治療前和治療后活檢樣本的比較,抗CTLA-4抗體治療后,Treg數量增加。因此,某些治療干預可能會增強Treg浸潤,從而影響后續治療的療效。
新的代謝機制
Tregs活化的核心是支持其生存和功能的脂質代謝的變化,這可由脂肪酸結合蛋白(FABPs)促進。此外,糖酵解和氧化代謝都有助于Tregs的擴增,因為腫瘤內Tregs在葡萄糖攝取方面的相對優勢可能促進FA合成。
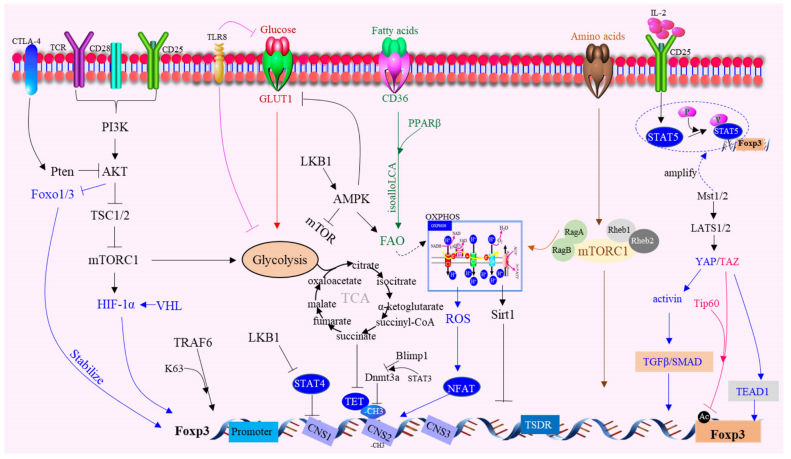
Tregs表現出另外一種獨特的代謝特征是線粒體代謝的增加。線粒體轉錄因子A(Tfam)和線粒體復合體III是維持Tregs免疫抑制功能調節基因表達所必需的。此外,Foxp3缺乏會導致mTORC2信號轉導的失調,并增強有氧糖酵解和氧化磷酸化。
值得注意的是,CD36在腫瘤內的Tregs中被選擇性上調,作為一種中樞代謝調節劑。CD36通過PPARβ信號精細調節線粒體適應性,重編程Tregs以適應富含乳酸的TME。在抗PD-1治療過程中,CD36靶向誘導了額外的抗腫瘤反應。
肝激酶B1(LKB1)調節Tregs的代謝和功能適應度,并作為DCs免疫原性的關鍵抑制劑發揮作用。在Tregs中,LKB1的特異性缺失可導致一種以Th2型顯性反應過多為特征的致命性炎癥性疾病,不僅破壞Tregs的生存、線粒體適應度和代謝,還可誘導免疫調節分子的異常表達,包括PD-1和TNF受體超家族蛋白的GITR和OX40。
新的遺傳機制
Helios可以增強人類胎兒CD4+幼稚T細胞向Tregs的優先分化,作為在炎癥反應期間穩定Tregs的關鍵轉錄因子。腫瘤中Helios缺陷的Tregs獲得效應T細胞功能,并通過上調對自身抗原具有高度親和力的效應細胞因子來促進對癌癥的免疫反應,如GITR/PD-1表達增加和對自身抗原的反應性增強。
Foxp1是一種forkhead轉錄因子,Foxp1的缺失會導致Tregs的功能受損,TSDR、CNS2、CNS3均為Foxp3相關的作用元件,調控著Foxp3的表達和功能。
YAP是Hippo通路的輔活化因子,在Tregs中高度表達,并促進Foxp3的表達和功能。TAZ是一種誘導Hippo信號轉導的TEAD轉錄因子的輔激活因子,它通過減少組蛋白乙酰轉移酶Tip60介導的Foxp3的乙酰化而減弱Tregs的發育。
核受體Nr4a是維持Treg基因程序的關鍵轉錄因子,參與Treg介導的TME抗腫瘤免疫抑制。促自噬蛋白AMBRA1也是T細胞的一個關鍵調節因子。此外,SUMO特異性蛋白酶3(SENP3)介導的BACH2去SUMO化阻止BACH2的核輸出,從而抑制與CD4+T效應細胞分化相關的基因,并穩定Treg特異性基因特征。
新的分子機制
在Tregs的分子機制方面也取得了一些新的進展。Tregs具有豐富的IL-2受體(IL-2R)表達,依賴于活化T細胞產生的IL-2,提示Tregs對IL-2的消耗與其抑制功能有關。并且IL-2R依賴的轉錄因子STAT5的激活在Tregs的抑制功能中起著重要作用。
絲氨酸-蘇氨酸激酶Mst1被鑒定為Tregs中IL-2-STAT5活性的信號依賴性放大器,Tregs中Mst1和Mst2的高活性對于防止腫瘤抵抗和自身免疫是至關重要的。
在T細胞活化過程中,Tregs中Foxp3的磷酸化可以通過TAK1 Nemo樣激酶(NLK)信號通路進行調節。另外,Tregs激酶TAK1的特異性缺失可以減少外周淋巴器官中Tregs的數量,而E3泛素化連接酶TRAF6缺乏的Tregs在體內功能紊亂。
ST2被認為是IL-33的唯一受體,表達ST2的Tregs對IL-33有反應,并且在IL-33刺激下Tregs的百分比增加,特別是Foxp3+GATA3+Tregs。在結直腸癌(CRC)的小鼠模型中,腫瘤浸潤的Tregs優先上調ST2,IL-33/ST2信號與腫瘤負荷呈正相關。
激活的Tregs表面表達糖蛋白-A為主的重復序列(GARP),它結合并激活潛伏的TGF-β。此外,整合素α4β1的激活也可增加Tregs的抑制能力。
Tregs穩態通過Tregs內在和外在機制與黏膜相關淋巴組織1(Malt1)功能密切相關。相比之下,CARD11-BCL10-MALT1(CBM)信號對于以Malt1蛋白酶依賴的方式介導Tregs的抑制功能至關重要。
抗CTLA-4單克隆抗體,如ipilimumab,通過Treg靶向改善TME的免疫特征。然而,在臨床研究中的詳細作用機制仍然存在爭議。在小鼠模型中,抗CTLA-4單克隆抗體誘導的抗腫瘤免疫反應完全依賴于通過Fc介導的ADCC對Treg的消耗。而在人類中,Treg的消耗與伊普利單抗抗腫瘤的主要療效無關。此外,靶向腫瘤內Tregs高表達分子的雙特異性抗體,如CTLA-4和OX-40或CTLA-4和GITR,它們可能通過有效地消耗體內的Treg來提高抗腫瘤療效。
由于腫瘤內Treg比其他效應T細胞表達更高水平的CD25,CD25可能是Treg耗竭的關鍵靶點。一些針對CD25的治療策略已經被FDA批準。例如,IL-2與白喉毒素的融合蛋白denileukin diftitox用于皮膚T細胞淋巴瘤。denileukin diftitox可與表達CD25的細胞結合,并通過白喉毒素的細胞毒性作用殺死它們。
新的抗CD25抗體RG6292被開發用于選擇性地清除Tregs,而不會干擾效應T細胞中的IL-2信號,目前正在進行臨床評估(NCT04158583)。靶向CD25的近紅外光免疫治療(NIR)也可能是腫瘤局部Treg去除的一種有希望的方法。此外,還開發了ADCT-301,一種抗CD25的抗體偶聯藥物,用于靶向表達CD25的淋巴瘤。它業可作為另一種在腫瘤中去除Tregs的選擇,目前正在進行臨床試驗(NCT03621982)。? ? ?
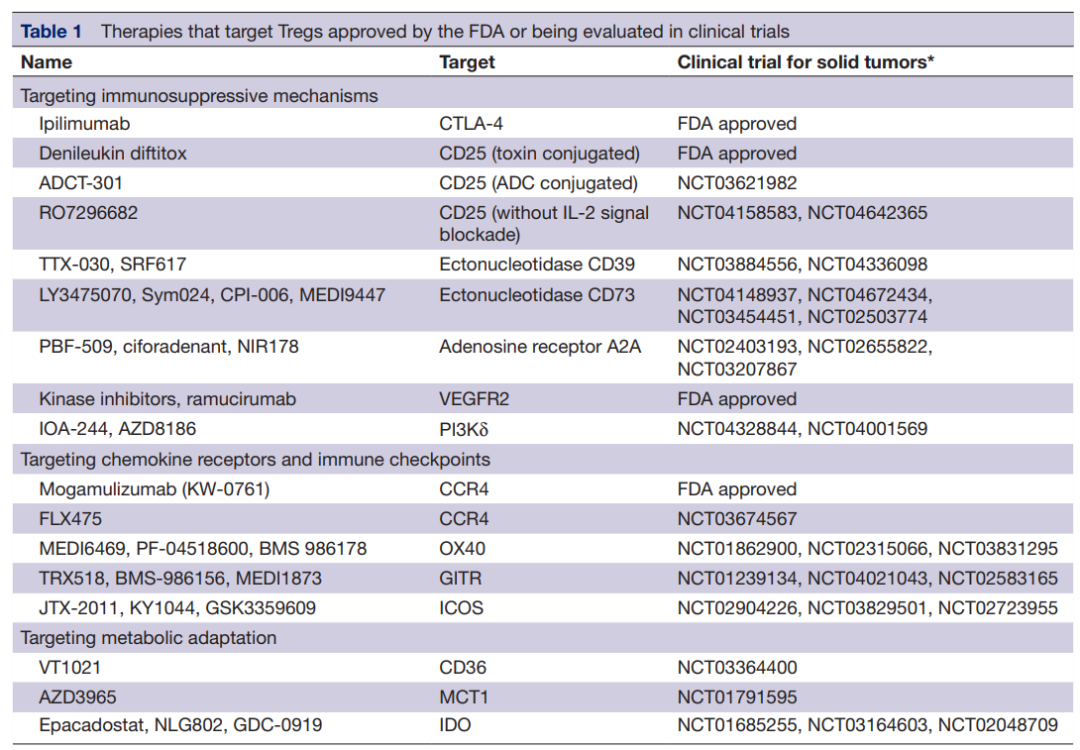
通過Tregs表達的外核苷酸酶CD39和CD73靶向腺苷途徑也可能是增強抗腫瘤免疫的一個有希望的靶點。阻斷外核苷酸酶活性的抗CD39和抗CD73抗體,如TTX-30、MEDI9447和BMS-986179,目前正在進行臨床試驗(NCT03884556、NCT03742102和NCT02754141)。除了抑制腺苷的產生外,腺苷受體A2AR的小分子抑制劑也可防止腺苷依賴性T細胞抑制。這些抑制劑的臨床試驗正在進行中(NCT02655822、NCT04089553和NCT02403193)。
此外,靶向VEGFA-VEGFR2軸可能通過減少TME中的Treg增殖和浸潤來激活抗腫瘤反應。已經證明,抗VEGFR2抗體ramucirumab可以減少胃癌患者eTreg的增殖。以FOXP3為靶點的療法也有開發。AZD8701是FOXP3的反義寡核苷酸,在體外實驗和人源化小鼠模型中部分降低了FOXP3及其下游轉錄分子的表達。AZD8701的臨床試驗正在進行中(NCT04504669)。
抗CCR4抗體mogamulizumab可減少實體瘤患者中CCR4+Treg的數量。在一項臨床試驗中,mogamulizumab聯合抗PD-1抗體(nivolumab)治療的患者中證實了Treg的缺失,這表明該組合是癌癥免疫治療組合中一個有希望的選擇。
由Treg高度表達的免疫檢查點分子,如OX40、GITR和ICOS,也可能成為治療靶點。一些研究表明,這些受體的刺激降低了Treg的免疫抑制功能,導致效應T細胞的激活。作為OX40激動劑的抗體,如MEDI6469134(NCT02274155),GITR,如MK-4166135(NCT02132754),以及ICOS,如KY1044136(NCT03829501),目前正在開展臨床研究。
在炎癥和非炎癥腫瘤中,TME中Treg浸潤、激活和存活的多種機制都已被揭示,影響TME中Treg浸潤、激活和存活的機制可能因患者而異。腫瘤細胞中的基因改變不僅決定免疫原性和炎癥狀態,而且也有助于調節TME中的免疫細胞浸潤和生存,例如基于趨化因子的Treg浸潤和基于代謝變化的Treg激活,這些都可以被特異性激酶抑制劑靶向。因此,TME中主要導致Treg浸潤的機制需要通過基于基因改變的免疫學和代謝譜來表征,并以精準免疫治療為目標。基于機制的Treg靶向治療有望改善目前的免疫治療。
參考文獻:
1.Mechanisms of regulatory T cellinfiltration in tumors: implications for innovative immune precision therapies.J Immunother Cancer.?2021; 9(7): e002591.
2. Regulatory T cells in tumor microenvironment: new mechanisms, potential
therapeutic strategies and future prospects. Mol Cancer. 2020; 19: 116.
3. Regulatory T Cells in Tumor Microenvironment and Approach for Anticancer Immunotherapy. Immune Netw. 2020 Feb; 20(1): e4.
?
編輯:小果果,轉載請注明出處:http://www.448371.com/cells/myxb/18036.html
免責聲明:本站所轉載文章來源于其他平臺,主要目的在于分享行業相關知識,傳遞當前最新資訊。圖片、文章版權均屬于原作者所有,如有侵權,請及時告知,我們會在24小時內刪除相關信息。
說明:本站所發布的案例均摘錄于文獻,僅用于科普干細胞與再生醫學相關知識,不作為醫療建議。
 微信掃一掃
微信掃一掃  支付寶掃一掃
支付寶掃一掃 