

一、微血管系統 — 二、基底膜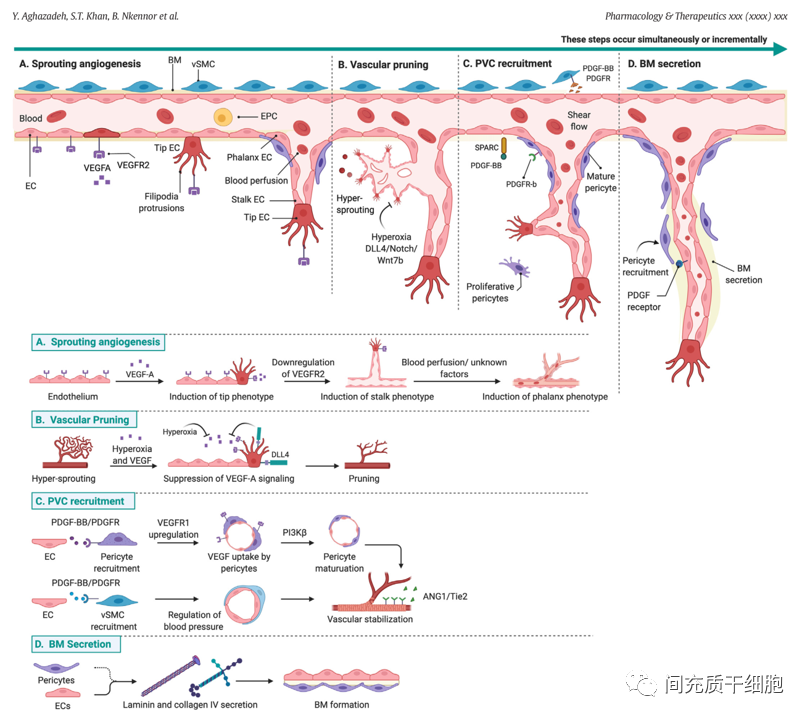

2.5被認為是冠狀動脈微血管功能障礙(CMD)[17]。皮膚微血管功能可以通過各種非侵襲性到輕度侵襲性的臨床評估來衡量[18]。
基底膜是血管系統的脫細胞成分,富含多種功能蛋白,由內皮細胞和血管周細胞分泌,對血管的完整性和穩定性至關重要。基底膜支持內皮細胞和血管周細胞之間的相互作用,并為血管和周圍非血管組織提供機械信號。基底膜已被鑒定出40多種基底膜蛋白質,它們的組合/比率在不同組織之間存在顯著差異[20]。
三、血管系統的關鍵細胞類型
(1)內皮細胞
血管周圍細胞的功能正常有助于血管完整性和屏障功能[24]。內皮細胞是血管周圍細胞群體中最重要的一種細胞類型。內皮細胞的α-平滑肌肌動蛋白(α-SMA)參與了血管收縮和血流調節。在靜止狀態下,內皮細胞也需要線粒體產生ATP的能量來維持生理功能和穩態。內皮細胞中大多數ATP是通過糖酵解代謝產生的,雖然內皮細胞的位置靠近血流并與足夠的氧氣接觸[25]。在內皮細胞糖酵解產生ATP的生理過程中,內皮細胞的生長和增殖伴隨著糖酵解代謝的增加,糖酵解代謝的增加又依賴于PFKFB3活性的上調,所以PFKFB3酶活性對調節內皮細胞的糖酵解至關重要[26-28]。
?
原代內皮細胞具有較高的免疫原性,異體移植治療能引起免疫排斥,因而需要搭配免疫抑制藥品的使用[29]。不過,大血管內皮細胞不表達MHC-II類分子,因此其免疫原性低于微血管內皮細胞[30]。再生研究中使用的血管細胞最常見的來源是來自人體組織的原代內皮細胞。原代人內皮細胞可從活組織或其他可獲得的組織中獲得,如臍帶靜脈和脂肪組織,并在體外擴增幾代。人臍靜脈內皮細胞(HUVEC)于1974年首次分離和培養[31],是臨床前研究中最常用的內皮細胞來源。因為內皮細胞根據血管結構樹的空間定位顯示出獨特的生理和轉錄組學特征[32, 33],因而人臍靜脈內皮細胞是從大血管靜脈獲得的,它們可能不適合生成微血管或動脈,因此,人臍靜脈內皮細胞和其他原代內皮細胞在臨床上的應用面臨著一些困難。
(2)周細胞(間充質干細胞)
周細胞嵌入血管基底層,包裹小直徑微血管,如毛細血管前小動脈、毛細血管和毛細血管后小靜脈。周細胞參與新生血管是通過內皮細胞和細胞外基質(ECM)的多種信號控制的,包括PDGF-BB和SPARC[39]。成熟周細胞通過表達VEGFR1來接收內皮細胞分泌的VEGF信號來調節血管生成,從而誘導內皮細胞出現莖表型并導致出芽方式的血管延長[40]。對成年小鼠的研究表明,周細胞數量的減少會導致出血和毛細血管過度擴張[41],并與糖尿病視網膜病變[42]和癌癥[43]等疾病相關。
(3)血管平滑肌細胞
在血管結構層次(動脈和靜脈、小動脈和小靜脈)中,血管平滑肌細胞(vSMC)是主要的血管周的壁細胞類型。血管平滑肌細胞呈梭形,通過細胞內豐富的αSMA來控制細胞的收縮功能,從而調節局部血壓[50, 51]。內皮細胞通過PDGF-BB信號與血管平滑肌細胞上的受體結合來吸引和黏附(招募)血管平滑肌細胞和周細胞[52, 53]。周細胞和血管平滑肌細胞分泌ANG1,激活內皮細胞表面的Tie-2受體,通過加強內皮細胞細胞間的相互作用來穩定血管;如果Tie受體出現突變,那么可以導致斑塊狀血管平滑肌細胞覆蓋和血管畸形,從而破壞血管的穩定性[54, 55]。
(4)其他細胞類型
除了周細胞和血管平滑肌細胞外,還有一些其他類型的細胞對內皮細胞起支持作用,如成纖維細胞[56]、中性粒細胞[57]和巨噬細胞[58]。在小鼠下肢缺血的實驗模型中,缺血周邊局部組織有多個明顯不同的成纖維細胞亞群(轉錄譜不同),而這些成纖維細胞亞群的存在有助于血管生成反應,而且還需要依賴于天然免疫信號的激活,才能有助于恢復缺血組織的灌注和恢復[59]。
四、血管病變的MSC治療
MSC通過分泌一些有功效的物質對受損組織發揮修復作用,或激活受損組織本身的干/祖細胞的激活和分化,即旁分泌作用機制,而不是MSC植入受損組織進行分化為組織細胞[60]。

—
(1)MSC的外泌體機制
MSC外泌體也可用于修復血管屏障功能,從而有利于治療血管屏障障礙相關的疾病[61]。在MSC外泌體的內含物中,miRNAs是非常重要的具有細胞生物學活性的作用物質之一。MiRNAs可作為基因表達的抑制子,與特定mRNA的3′-非翻譯(3′-UTR)區域結合,可通過促進mRNA降解或抑制mRNA翻譯為蛋白質,在共同途徑內同時調節多個基因[62, 63]。

—
MSC分泌的外泌體中,同時存在促進血管生成的miRNAs和抑制血管生成的miRNAs,不過抑制血管生成的miRNAs含量很低[64]。比如,miR-93通過作用于多個調節基因,增強內皮細胞增殖和減少骨骼肌細胞凋亡,從而促進后肢缺血后的毛細血管密度和灌注恢復[65]。外泌體所含的miR-125a可通過靶向抑制delta-like 4(DLL4)的表達,從而解除了delta-like 4對血管生成的抑制作用,進而促進內皮細胞的血管生成[66]。又如,miR-15a[67]和miR-503[68]同樣為內皮細胞增殖和血管生成的負性調節因子。miR-92a和miR-100靶向對應于幾種促血管生成蛋白的mRNAs,在肢體缺血和心肌梗死的小鼠模型中,抑制miR-19[69]、miR-92a[70]和miR-100[71]的活性可促進血管生長和受損組織的功能恢復。抑制可改善老齡小鼠缺血后的血流恢復,并減少對這些靶點的抑制。
(2)MSC的抗氧化作用
MSC介導的血管再生的一個主要因素是宿主組織的抗氧化作用[77, 78]。MSC對氧化應激具有高度抵抗力,因為它們組成性地表達過氧化氫酶、超氧化物歧化酶1-3(SOD1-3)、谷胱甘肽過氧化物酶(GPx)、SIRT 1/3/6,以及硫氧還蛋白、血紅素加氧酶-1,谷胱甘肽(GSH)。這些抗氧化物質通過降低氧化應激、減少炎癥以及增加宿主內皮細胞/血管平滑肌細胞抗氧化蛋白含量和/或活性,介導血管損傷和疾病的恢復[79-81]。
(3)MSC的細胞因子的作用
在MSC的培養上清中,MSC分泌多種可溶性因子-包括VEGF、VEGFR2、Ang-1/Tie2、單核細胞趨化蛋白-1(MCP-1)、巨噬細胞炎性蛋白-1α(MIP-1α)、MIP-1β、單因子、堿性成纖維細胞生長因子(bFGF)、PDGF、IL-1β、IL-10、干細胞衍生因子(SDF)-1、肝細胞生長因子(HGF)、胰島素樣生長因子1(IGF-1)、胸腺肽-β4、Wnt5a 等[60, 84]。
—
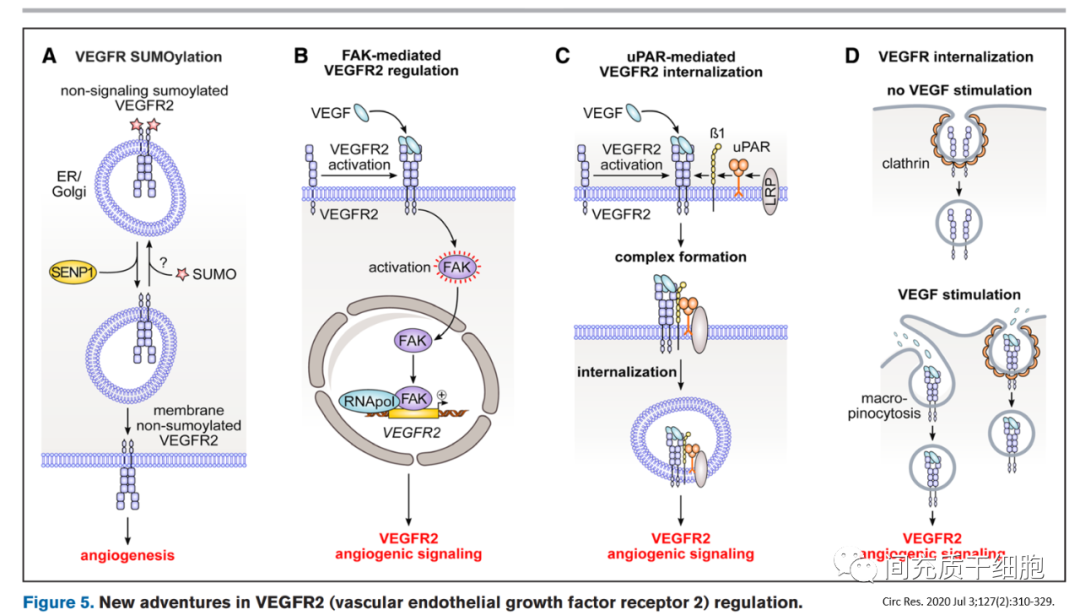
在缺氧和血清饑餓狀態下,向內皮細胞中添加VEGF165a可通過減少細胞死亡和增加內皮細胞增殖以及增加糖酵解來誘導血管生成,但存在血管通透性和血管滲漏的現象[89]。而且人體臨床研究使用VEGF165a等藥物促進缺血區域血管生長以重建血供的治療效果不佳[90]。
(4)其他機制
MSC來源的外泌體能改善受損細胞的線粒體功能,減少線粒體DNA的損傷和提高線粒體NDA的穩定性[91-93]。甚至在內皮細胞發生缺血/再灌注損傷的實驗中,可以觀察到MSC的線粒體可以直接從隧道納米管轉移給內皮細胞,增加內皮細胞產生更多的能量ATP,從而挽救損傷的內皮細胞[94, 95]。在脂多糖誘導的急性肺損傷模型中,肺泡上皮細胞活性氧增加、線粒體膜電位降低和ATP產生降低,肺部血管屏障功能減弱;MSC與肺泡上皮形成了含有連接蛋白43(Cx43)的縫隙連接通道,釋放出被上皮細胞吞噬的含有線粒體的外囊泡(外泌體),線粒體從MSC轉移到肺泡上皮細胞導致肺泡ATP濃度增加,減少了LPS誘導的急性肺損傷炎癥[96]。
五、小結
MSC治療也可以在高血壓和外周阻力增加的情況下促進微血管再生。比如在慢性腎動脈狹窄大鼠模型中,MSC治療促進血管緊張素2型受體表達增加、恢復狹窄腎臟的微血管樹狀網絡結構,從而提高腎小球濾過率和腎血漿流量[100]。MSC能恢復肝臟微血管功能和降低門靜脈高壓力[101]。然而,自體干細胞療法的持續給藥受到患者可獲取數量的限制,以及捐贈或接受細胞療法的患者是否存在可能影響細胞療法的病理狀況[102]。
it is naive to think that one administration of cells is enough to provide
life-long therapeutic remedy.)

—
簡而言之,在基于細胞的再生醫學中,想在體內重建成熟的具有功能的網狀結構血管,仍然是有待突破的科學難題。單細胞表觀基因組學和轉錄組學的檢測技術有可能分析細胞隨時間變化的軌跡,這無疑將有助于確定血管再生過程中血管成熟的關鍵細胞生物學變化。相信隨著對血管再生各個環節的深刻理解,借助于MSC和多種細胞的相互作用,將來是可以完美地解決血管生成的難題。
?
-END-
?
來源:間充質干細胞
編輯:小果果,轉載請注明出處:http://www.448371.com/cells/gxb/33668.html
免責聲明:本站所轉載文章來源于其他平臺,主要目的在于分享行業相關知識,傳遞當前最新資訊。圖片、文章版權均屬于原作者所有,如有侵權,請及時告知,我們會在24小時內刪除相關信息。
說明:本站所發布的案例均摘錄于文獻,僅用于科普干細胞與再生醫學相關知識,不作為醫療建議。
 微信掃一掃
微信掃一掃  支付寶掃一掃
支付寶掃一掃 