

前言
癌基因及其前體——原癌基因的發(fā)現(xiàn)促使人們提出了一系列問題,其中居于核心地位的問題是,癌基因到底如何通過它們所編碼的蛋白質(zhì)成功地擾亂了細胞的行為?
對這個問題的理解,首先要了解正常細胞的生命活動。細胞接收并處理生化信號從而調(diào)節(jié)細胞增殖,正常的癌基因編碼蛋白質(zhì)常常是這一機制的組成部分。正常組織形態(tài)維持的實現(xiàn)皆有賴于細胞間的協(xié)同合作,而大部分對話是由生長因子負責(zé)傳達的。這些相對分子量較小的蛋白經(jīng)細胞分泌,于細胞間隙運行,最終作用于其他細胞的受體,傳遞它們獨特的生物學(xué)信息。
在癌癥中,使細胞感知周圍環(huán)境中生長因子的存在、將信號傳入細胞內(nèi)部,并進一步處理這些信號的信號分子被癌基因編碼蛋白質(zhì)篡權(quán)。通過取代正常細胞的生長刺激機制,癌蛋白可誘使細胞相信周圍環(huán)境中存在的正是生長所需的生長因子。一旦誘導(dǎo)成功,細胞便會開始增殖,其反應(yīng)與大量生長因子存在于其周圍介質(zhì)所介導(dǎo)的反應(yīng)一樣。
?
癌蛋白和生長因子受體的
酪氨酸激酶功能
細胞間信號如何通過生長因子傳遞的第一條線索來源于對癌基因v-src及其特定蛋白質(zhì)產(chǎn)物的生化分析。1978年,生化學(xué)家發(fā)現(xiàn)了Src發(fā)揮作用的機制。他們研制出能與Src特異識別并結(jié)合的抗體,當(dāng)其在同時含有Src及腺苷三磷酸的溶液中孵育時,該抗體被磷酸化。由此可知,Src是作為蛋白激酶發(fā)揮作用的,即將ATP的一個高能磷酸鍵轉(zhuǎn)移至一個適宜的蛋白質(zhì)底物。? ? ? ? ? ? ? ? ?
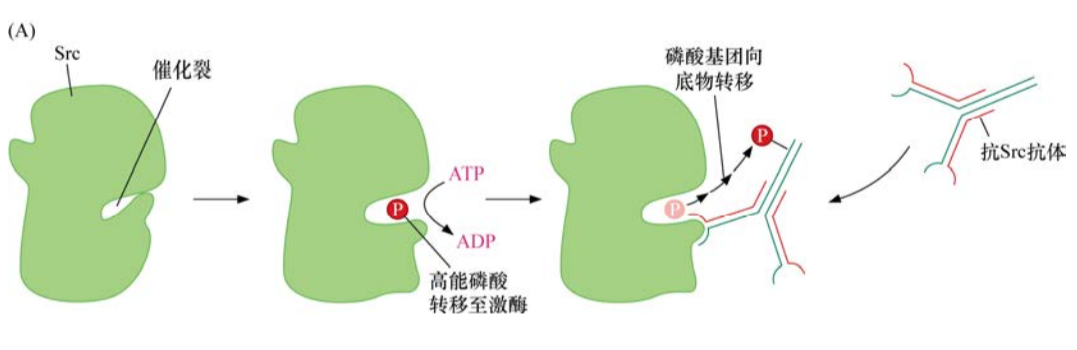

Src激酶功能的揭示意義深遠。原則上,一個蛋白激酶可以磷酸化細胞內(nèi)多種不同的蛋白質(zhì)底物。一旦被磷酸化,這些底物蛋白質(zhì)的功能可能會發(fā)生改變,進而改變其下游靶點的功能。這種作用機制似乎可以解釋像Src這樣的蛋白為何能如此多效地干擾多種細胞的表型。
不久之后,人們又發(fā)現(xiàn)Src與以前發(fā)現(xiàn)的其他蛋白激酶顯著不同。已知其他的蛋白激酶是將磷酸基團連接到絲氨酸和蘇氨酸殘基的側(cè)鏈上。而然Src則磷酸化蛋白質(zhì)底物的酪氨酸殘基。在正常細胞內(nèi)99%以上的磷酸化氨基酸為磷酸化絲氨酸和磷酸化蘇氨酸,磷酸化酪氨酸僅占這些細胞全部磷酸化氨基酸的0.05%-0.1%。
在隨后的研究中人們得出了另一條與之相關(guān)的結(jié)論:在哺乳動物細胞中,通過酪氨酸磷酸化傳遞信號的這種機制主要出現(xiàn)在絲裂原信號轉(zhuǎn)導(dǎo)通路中,而其他信號轉(zhuǎn)導(dǎo)途徑中所涉及的成千上萬的激酶,則基本上全部依賴于磷酸化絲氨酸和磷酸化蘇氨酸來傳遞信息。那么酪氨酸殘基的磷酸化是如何導(dǎo)致細胞轉(zhuǎn)化的呢?
對這一問題的回答要提到上皮生長因子——EGF。人們發(fā)現(xiàn),EGF對多種上皮細胞具有促有絲分裂作用。EGF通過作用于一種細胞表明蛋白——EGF受體(EGFR),并與之結(jié)合,將信息向胞內(nèi)傳遞。對EGFR的整體結(jié)構(gòu)特征分析,發(fā)現(xiàn)EGFR分為3個主要區(qū)域,較大的N端胞外結(jié)構(gòu)域,主要負責(zé)識別并結(jié)合EGF;一段23個氨基酸的跨膜區(qū),這段疏水氨基酸構(gòu)成區(qū)域可以在細胞膜的脂質(zhì)雙層中穩(wěn)定存在;最后,C端由542個氨基酸構(gòu)成的第三個結(jié)構(gòu)域,伸展進入細胞質(zhì)。對胞內(nèi)結(jié)構(gòu)域的檢測顯示,它與已知的Src蛋白質(zhì)序列有明確的序列相似性。
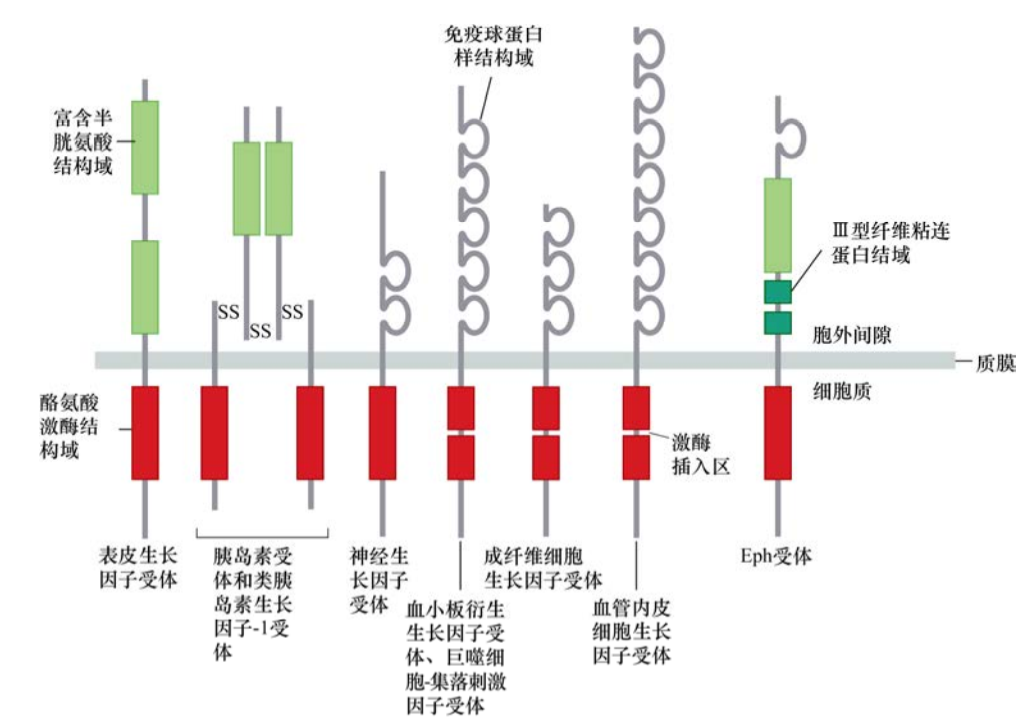
這樣,答案瞬間變得清晰起來:一旦EGFR結(jié)合EGF,其胞內(nèi)結(jié)構(gòu)域的Src樣激酶將以某種方式被激活,之后磷酸化某些胞內(nèi)蛋白質(zhì)的酪氨酸殘基,從而使細胞增殖。隨后的測序試驗結(jié)果顯示,許多酪氨酸激酶都有整體的結(jié)構(gòu)相似性,其中許多可產(chǎn)生癌蛋白樣作用。
被改變的生長因子受體可發(fā)揮
癌蛋白樣作用
1984年,人們證實EGF受體序列與已知的由erbB癌基因編碼的癌蛋白序列高度相關(guān),這一發(fā)現(xiàn)極具震撼力。由此,細胞生物學(xué)的兩個不同領(lǐng)域被聯(lián)系起來,一個被細胞用于感知周圍環(huán)境中生長因子的蛋白質(zhì)被挪用為一種由逆轉(zhuǎn)錄病毒基因編碼的強力癌蛋白。
一次具體的研究中,人們發(fā)現(xiàn)erbB癌基因編碼的癌蛋白缺少EGF受體N端胞外結(jié)構(gòu)域的序列。在沒有N端序列的情況下,ErbB癌蛋白無法辨認(rèn)及結(jié)合EGF,然而,它發(fā)揮了細胞增殖刺激因子的功效。這一發(fā)現(xiàn)引發(fā)了一個有趣的推論并隨后被證實:胞外結(jié)構(gòu)域的缺失可通過某種方式使被截短的EGF受體蛋白向細胞內(nèi)源源不斷地發(fā)送生長刺激信號,而完全不依賴于EGF。多年后,人們在1/3的人成膠質(zhì)細胞瘤中發(fā)現(xiàn)了這些被截短的EGF受體。而后,在乳腺癌中發(fā)現(xiàn)了ErbB/EGF-R的同源分子,如ErbB2、HER2及Neu,它們與不良預(yù)后密切相關(guān)。
ErbB-EGFR之間的關(guān)系為癌細胞的特性提供了一個簡單明了的解釋:ErbB癌蛋白所釋放的信號與那些經(jīng)配體激活的EGF受體所釋放的信號非常相似。然而,不同于EGFR的是,ErbB癌蛋白可以持續(xù)不斷地向細胞內(nèi)發(fā)放生長刺激信號,從而使細胞認(rèn)為周圍存在著大量的EGF,而其實那里可能什么都沒有。
生長因子基因可轉(zhuǎn)變?yōu)榘┗?/strong>
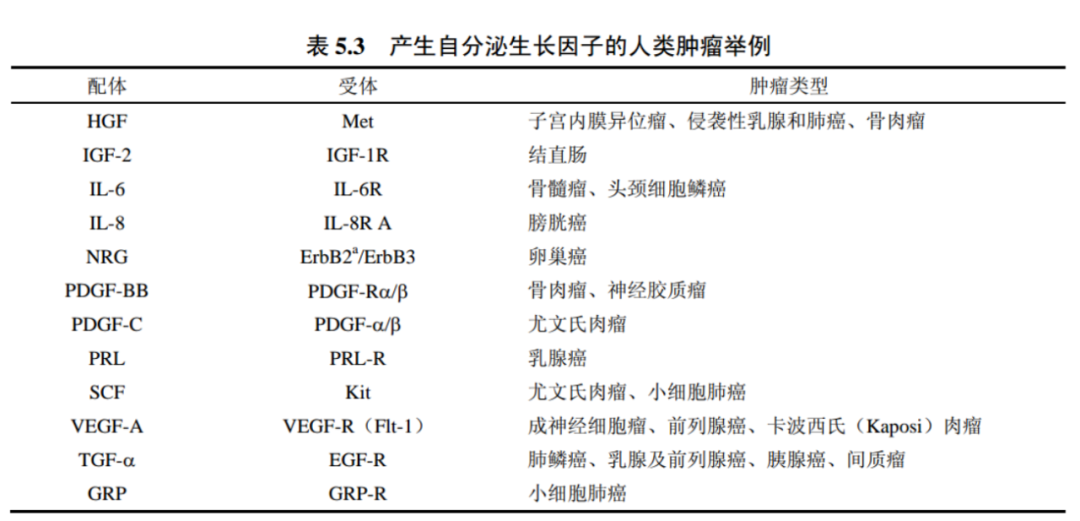
1983年,人們發(fā)現(xiàn)PDGF的B鏈序列與由猴肉瘤病毒v-sis癌基因編碼的癌蛋白高度相關(guān)。PDGF與sis-編碼癌蛋白間的聯(lián)系向我們提供了另一個癌蛋白轉(zhuǎn)化細胞的重要機制:猴肉瘤病毒感染細胞后,它的sis癌基因促使被感染細胞釋放大量PDGF樣sis蛋白到其周圍細胞外空間中。在那里,PDGF樣分子與釋放它們的自身細胞上的PDGFR結(jié)合。其結(jié)果是造成這些細胞上PDGF受體強烈活化,在大量細胞中該配體激活PDGFR產(chǎn)生持續(xù)的生長刺激信號。
有絲分裂信號體系中的蛋白質(zhì)又一次與病毒癌蛋白緊密地聯(lián)系在了一起。病毒感染細胞合成并釋放刺激自身產(chǎn)生應(yīng)答的生長因子,而并非由一種類型的細胞向其附近的另一種類型的細胞發(fā)送生長因子信號(旁分泌),或由機體中的一種組織經(jīng)由循環(huán)系統(tǒng)向另一遠隔組織發(fā)放信號(內(nèi)分泌)。它代表的是一種自我刺激,或自分泌信號系統(tǒng),該系統(tǒng)中的細胞可生產(chǎn)作用于自身的有絲分裂原。
事實上,我們已知許多腫瘤細胞可以產(chǎn)生并釋放大量它們自身亦可應(yīng)答的生長因子。
受體酪氨酸激酶依賴磷酸化發(fā)揮作用
通過為細胞提供持續(xù)生長信號刺激,癌蛋白能夠驅(qū)動周而復(fù)始的細胞生長、分裂,那么具有酪氨酸激酶活性的生長因子受體是如何成功地將信號從胞外傳遞到胞內(nèi)的呢?
為了解決這個問題,研究者運用EGF生長因子處理能夠表達其同源受體EGFR的細胞,幾秒鐘后檢測蛋白的磷酸化。結(jié)果顯示,伴隨配體與生長因子受體的結(jié)合,多種胞質(zhì)蛋白的酪氨酸殘基發(fā)生了磷酸化。這些蛋白中發(fā)生酪氨酸磷酸化最顯著的是受體分子本身,因此,這些受體看起來是能夠自磷酸化的。

另一個線索來源于很多生長因子配體的結(jié)構(gòu):它們通常是二聚體,形成同源二聚體或異源二聚體。第三條線索來源于很多跨膜蛋白有類似EGF和PDGF受體的結(jié)構(gòu),在質(zhì)膜上具有橫向流動性。這些線索提供了一個簡單的模型,當(dāng)沒有配體的情況下,一個生長因子受體通常以單體的形式嵌入質(zhì)膜。當(dāng)其生長因子配體存在時,受體分子就會結(jié)合在配體兩個亞基的一個上。其后,配體-受體復(fù)合物將在質(zhì)膜上徘徊直到有第二個機會遇到另一個受體分子,配體中沒有結(jié)合受體的那個亞基將和第二個受體分子結(jié)合。這樣,通過二聚體形式的配體為橋梁,把兩個受體分子成功地交聯(lián)在一起。
?
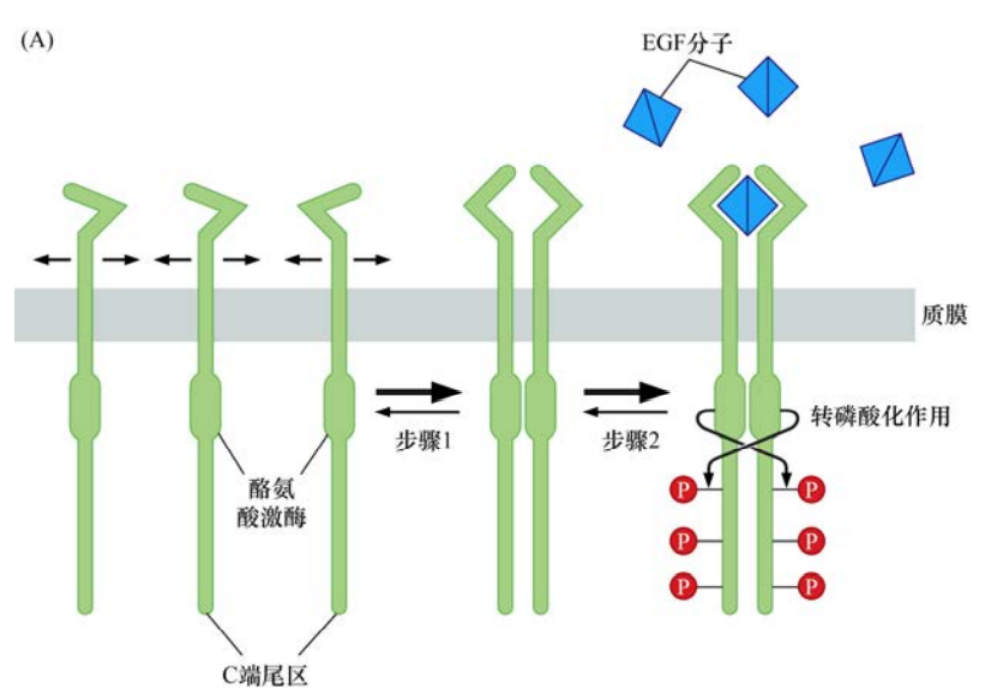
一旦兩個受體分子的胞外結(jié)構(gòu)域通過與配體結(jié)合而發(fā)生二聚化,胞質(zhì)部分通常也會被拉在一起。每個激酶結(jié)構(gòu)域磷酸化另一個受體胞質(zhì)結(jié)構(gòu)域里的酪氨酸殘基。這種雙向的相互磷酸化作用被稱為轉(zhuǎn)磷酸作用。
這些酪氨酸殘基的磷酸化至少導(dǎo)致兩種后果。激酶催化裂隙(即蛋白酶發(fā)揮功能的區(qū)域)通常受蛋白質(zhì)的環(huán)結(jié)構(gòu)阻隔,從而阻止激酶與其底物有效結(jié)合。阻隔“活化環(huán)”的關(guān)鍵酪氨酸的轉(zhuǎn)磷酸作用可導(dǎo)致該環(huán)結(jié)構(gòu)移開,從而可使底物分子直接進入催化裂隙。另外,轉(zhuǎn)磷酸作用還可導(dǎo)致生長因子受體胞質(zhì)部分激酶區(qū)域以外的一系列酪氨酸殘基的磷酸化。這些磷酸化事件可使受體分子激活多種下游信號通路。
編輯:小果果,轉(zhuǎn)載請注明出處:http://www.448371.com/linchuang/lcyj/23448.html
免責(zé)聲明:本站所轉(zhuǎn)載文章來源于其他平臺,主要目的在于分享行業(yè)相關(guān)知識,傳遞當(dāng)前最新資訊。圖片、文章版權(quán)均屬于原作者所有,如有侵權(quán),請及時告知,我們會在24小時內(nèi)刪除相關(guān)信息。
說明:本站所發(fā)布的案例均摘錄于文獻,僅用于科普干細胞與再生醫(yī)學(xué)相關(guān)知識,不作為醫(yī)療建議。
 微信掃一掃
微信掃一掃  支付寶掃一掃
支付寶掃一掃 