▉ 摘要
T細(xì)胞 受體、共刺激信號和細(xì)胞因子 信號協(xié)調(diào)地支配觸發(fā) T細(xì)胞激活和功能編程 的特定信號網(wǎng)絡(luò)。了解 免疫代謝信號網(wǎng)絡(luò) 的 調(diào)節(jié)器和效應(yīng)器 可能會發(fā)現(xiàn)調(diào)節(jié)人類疾病中代謝程序和T細(xì)胞反應(yīng)的治療靶點(diǎn) 。
在這篇文章中,我們總結(jié)了 絲氨酸/蘇氨酸激酶 介導(dǎo)的關(guān)鍵信號網(wǎng)絡(luò)的上游調(diào)節(jié)器 和信號效應(yīng)器 ,包括調(diào)節(jié)代謝的 PI3K–AGC激酶 、 mTOR 和 LKB1–AMPK 途徑,特別是在 T細(xì)胞中的調(diào)節(jié)作用 。我們重點(diǎn)討論了參與免疫代謝調(diào)節(jié)的細(xì)胞信號通路,主要涉及參與代謝調(diào)節(jié)的相關(guān)分子、上游和下游靶點(diǎn)及其細(xì)胞類型特異性效應(yīng) ,分為PI3K、AGC、mTOR的機(jī)制靶點(diǎn)和LKB1–AMPK信號 。
當(dāng)T細(xì)胞受體 (TCR) 在 共刺激和細(xì)胞因子 信號 存在的情況下識別同源抗原時,原始T細(xì)胞中的信號網(wǎng)絡(luò)被 激活 ,以促進(jìn) 克隆擴(kuò)增 、效應(yīng)細(xì)胞分化和免疫功能。在抗原清除后,大多數(shù)效應(yīng)細(xì)胞通過 程序性細(xì)胞死亡 而死亡,但一些細(xì)胞作為 記憶 細(xì)胞則會長期存在,并為 快速回憶 進(jìn)行下一次免疫斗爭做好準(zhǔn)備。此外,T細(xì)胞的功能狀態(tài)有著相同和不同的 轉(zhuǎn)錄程序 ,以及 蛋白質(zhì)表達(dá)、活性和相互作用的差異 。在現(xiàn)階段的研究中,這些差異才剛剛開始被理解,人們也越來越認(rèn)識到細(xì)胞代謝在決定T細(xì)胞發(fā)育和功能方面的重要性。
▉1.? PI3K-AGC signaling
磷脂 是影響下游免疫代謝途徑的關(guān)鍵第二信使,因此在轉(zhuǎn)換方面受到高度調(diào)節(jié)。磷脂轉(zhuǎn)換的一個 中心介質(zhì) 是 PI3K ,它將磷脂酰肌醇-(4,5)-二磷酸 (PIP2) 轉(zhuǎn)化為磷脂酰肌醇-(3,4,5)-三磷酸 (PIP3) 。 PIP3 的產(chǎn)生引起質(zhì)膜募集 和含有PH (pleckstrin homology) 結(jié)構(gòu)域的蛋白質(zhì)功能調(diào)節(jié) 。
?
因此,PI3K活性可以通過將大量 含有PH結(jié)構(gòu)域的效應(yīng)蛋白招募 到鄰近區(qū)域來 產(chǎn)生亞細(xì)胞信號中樞 。PI3K活性誘導(dǎo)參與調(diào)節(jié)細(xì)胞功能的多種信號通路,包括 Akt (蛋白激酶B) 、磷脂酰肌醇依賴性蛋白激酶1 (PDK1) 以及mTOR復(fù)合物1 。
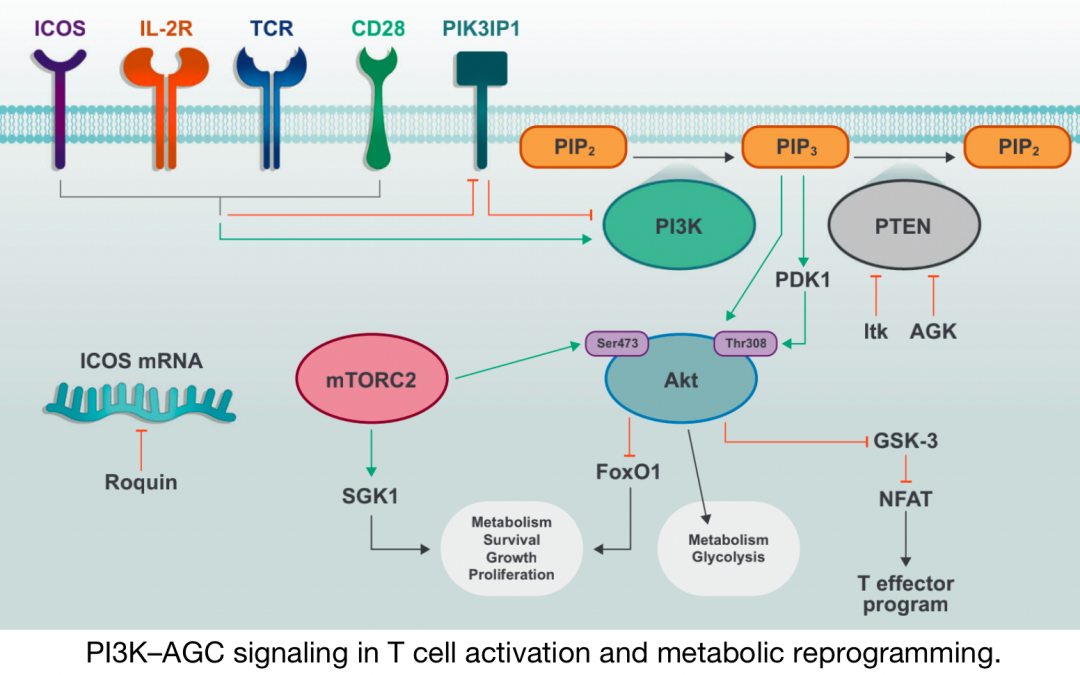
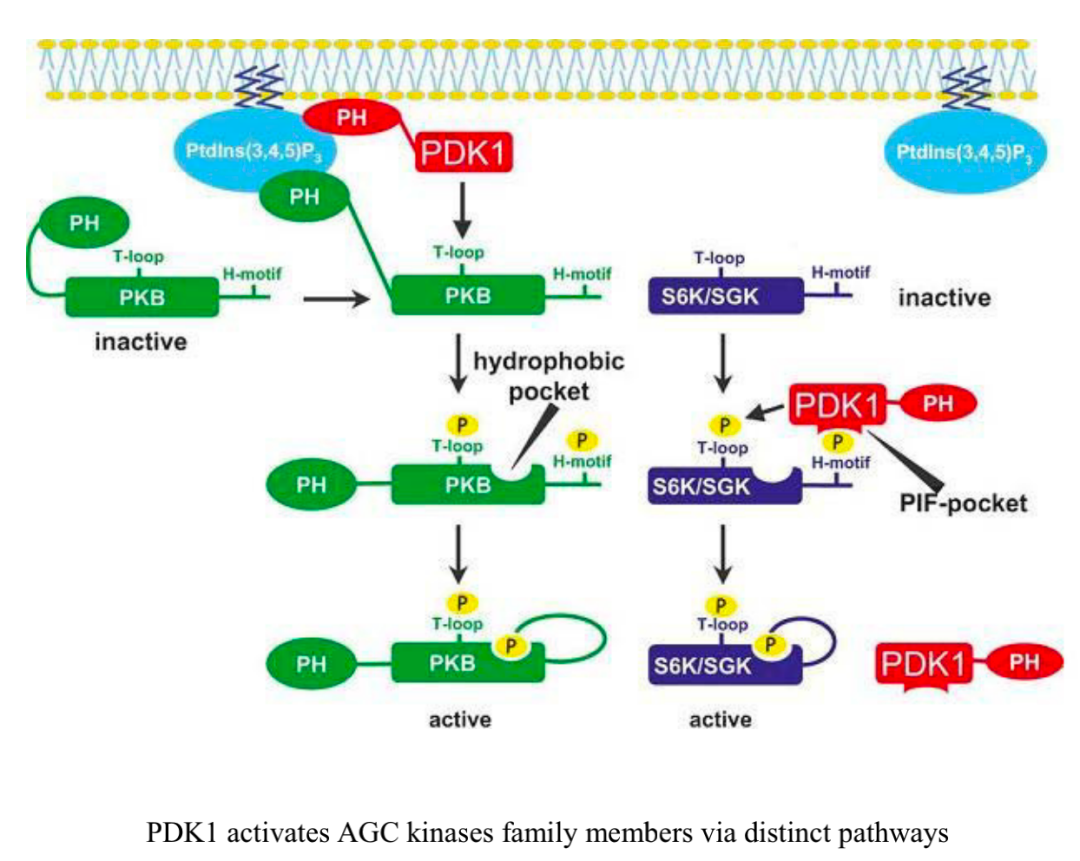
PI3K–T細(xì)胞活化和代謝重編程中的AGC信號
TCR、CD28和IL-2R 的激活誘導(dǎo) PI3K 的磷酸化和激活 ,以及PI3K抑制分子的失活 。PIP2通過PI3K 的活性轉(zhuǎn)化為PIP3 ,PIP3促進(jìn)包括 PDK1 和 Akt 在內(nèi)的下游信號分子的質(zhì)膜募集和激活 。 mTORC2 進(jìn)一步激活A(yù)kt并促進(jìn)代謝增加 和T細(xì)胞效應(yīng)器功能 。
▉1.1 PI3K
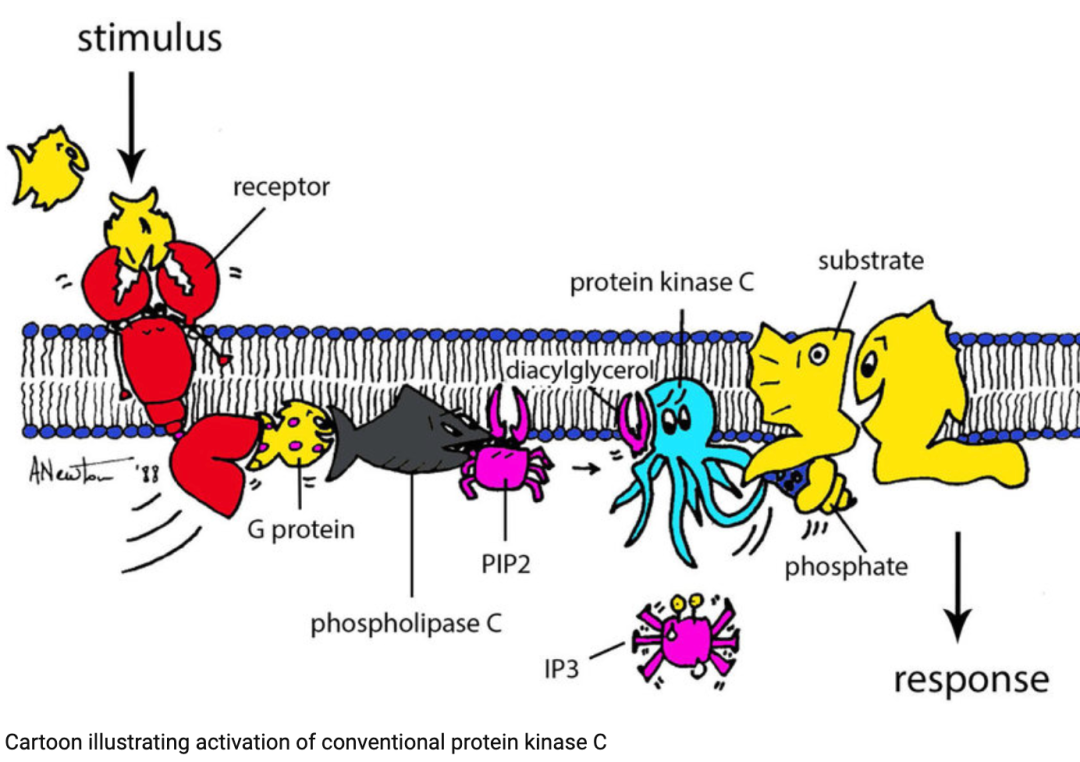
《Protein kinase C: perfectly balanced》
I類 PI3K 是一種異二聚體蛋白質(zhì) ,由一個催化的p110亞單位 (p110α、p110β、p110δ或p110γ) 和五個調(diào)節(jié)亞單位亞型 (p50α, p55α, p55γ, p85α, or p85β) 中的一個組成。在沒有刺激 的情況下, 催化亞單位 的激酶結(jié)構(gòu)域 被調(diào)節(jié)亞單位 抑制 。PI3K底物的濃度與細(xì)胞激活狀態(tài)密切相關(guān),在T細(xì)胞中,細(xì)胞激活狀態(tài) 由TCR結(jié)合、CD28家族介導(dǎo)的共刺激和細(xì)胞因子信號 (如IL-2R下游) 決定。在上游激活 后,PI3K的Src-homology-2 (SH2) 結(jié)構(gòu)域可結(jié)合上游受體或銜接蛋白 上的特異性磷酸化YXXM基序 ,導(dǎo)致調(diào)節(jié)亞單位的抑制性連接釋放和催化亞單位移位至富含底物的質(zhì)膜 。
?
PI3K信號由 磷酸酶活性負(fù)調(diào)控 。具體而言,PIP3分別通過磷酸酶 和張力蛋白 同系物 (PTEN) 以及含SH2結(jié)構(gòu)域 的肌醇5′-磷酸酶 (SHIP) 轉(zhuǎn)化為PIP2 或PI-(3,4)-P2 。SHIP在促進(jìn)Th1細(xì)胞反應(yīng) 中起著至關(guān)重要的作用,而PTEN缺乏 會導(dǎo)致T細(xì)胞過度活化,尤其是在次優(yōu)刺激下。PTEN在強(qiáng)TCR刺激或CD28共刺激下被抑制 ,部分原因是Tec家族激酶IL-2誘導(dǎo) 的T細(xì)胞激酶 (Itk) ,并且在CD8 T細(xì)胞激活期間也受到AGK介導(dǎo)的磷酸化的負(fù)調(diào)節(jié) 。
在最顯著的 含有PH結(jié)構(gòu)域 的蛋白質(zhì)中有 AGC激酶 的成員,包括PDK1、Akt、核糖體S6激酶 (RSK,也稱為p90) 和血清/糖皮質(zhì)激素調(diào)節(jié)激酶1 (SGK1) 。 PDK1 是一種絲氨酸/蘇氨酸激酶,在激活其他AGC激酶 (包括Akt和SGK1) 中起重要作用。TCR和CD28激活促進(jìn)PDK1向質(zhì)膜的募集并誘導(dǎo)其磷酸化 , Thr513處 PDK1的蛋白激酶C-θ (PKC-θ) 依賴性磷酸 化, 而不是激酶結(jié)構(gòu)域殘基 (Ser241) 的自動磷酸化 ,這對于 驅(qū)動T細(xì)胞活化 至關(guān)重要。在活化的CD8 T細(xì)胞中, PDK1參與代謝重編程 ,維持IL-2刺激下游的葡萄糖攝取和糖酵解 ,該過程需要mTOR–HIF-1α (缺氧誘導(dǎo)因子-1α) 軸 ,但不需要PI3K或Akt活性。PDK1活性的適當(dāng)調(diào)節(jié)對于 加強(qiáng)適當(dāng)?shù)腡細(xì)胞活化以及控制炎癥的Treg細(xì)胞功能 至關(guān)重要。在Ser473處的mTORC2磷酸化后,Akt 允許 Akt結(jié)合到被稱為PDK1的“PIF口袋”的底物dock ,這促進(jìn)了Thr308處Akt的磷酸化 。 mTORC2–Akt 信號則可以協(xié)調(diào)T細(xì)胞激活、分化和運(yùn)輸 。
《 The Clinical Implications of the Survival Pathway in Prostate Cancer 》
Akt在T細(xì)胞中的一個重要作用是通過磷酸化調(diào)節(jié)叉頭盒O (FoxO) 轉(zhuǎn)錄因子 的活性,這導(dǎo)致它們被排除在細(xì)胞核之外并終止靶基因轉(zhuǎn)錄 。 FoxO蛋白 在靜止的細(xì)胞群中更具轉(zhuǎn)錄活性,它們通過KLF2促進(jìn)穩(wěn)態(tài)細(xì)胞因子受體IL7R的表達(dá)以及細(xì)胞運(yùn)輸分子 (如CD62L、CCR7和S1PR1) 的表達(dá)。最近的證據(jù)表明FoxO1活性的適當(dāng)下調(diào)對T細(xì)胞內(nèi)環(huán)境穩(wěn)定至關(guān)重要。從機(jī)制上講,活化T細(xì)胞中的FoxO1下調(diào)對于協(xié)調(diào)細(xì)胞生長和增殖非常重要,因?yàn)樗试S持續(xù)的mTORC1信號和合成代謝,因此, Akt–FoxO1軸動態(tài)調(diào)節(jié)T細(xì)胞反應(yīng) 。
?
另一種參與Akt信號傳導(dǎo)的絲氨酸/蘇氨酸激酶是 糖原合成酶激酶3 (GSK-3) ,它在靜息細(xì)胞 中具有組成性活性,但通過Akt的磷酸化而失活。活性GSK-3在T和B細(xì)胞中通過限制活化T細(xì)胞的核因子 (NFAT) 活性,促進(jìn)其存活并限制活化。AGC激酶 (如RSK) 及其底物在T細(xì)胞代謝規(guī)劃 中的確切作用一直備受關(guān)注,也同樣是目前研究的重點(diǎn)方向。
▉2.? mTOR signaling
mTOR 是一種絲氨酸/蘇氨酸蛋白激酶,存在于兩種信號復(fù)合物 mTORC1 和 mTORC2 中 。mTORC1由mTOR激酶定義,mTORC1的適配蛋白調(diào)節(jié)相關(guān)蛋白和SEC13蛋白8 (MLST8) 的哺乳動物致死蛋白,以及脯氨酸Akt底物-1 (PRAS1) 及其靶DEP結(jié)構(gòu)域相互作用蛋白 (DEPTOR) 的抑制亞單位。 m TORC1信號對胸腺中的T細(xì)胞發(fā)育、外周的內(nèi)環(huán)境穩(wěn)定和分化 為效應(yīng)CD4 Th1、Th2和Th17細(xì)胞以及細(xì)胞毒性CD8 T細(xì)胞至關(guān)重要。 相比之下,Th1和Th2細(xì)胞分化 更需要mTORC2活性,同時后者也能調(diào)節(jié)Tfh和Treg細(xì)胞的遷移 。
?
mTOR信號通過調(diào)節(jié)體內(nèi) Treg細(xì)胞的激活、譜系穩(wěn)定性和抑制功能 ,在對抗傳統(tǒng)T細(xì)胞反應(yīng)中發(fā)揮額外作用 。最后,mTOR信號可拮抗淋巴組織中 長期記憶CD8 T細(xì)胞的分化 ,卻可促進(jìn)其在非淋巴組織中的發(fā)育 。因此,mTOR信號是T細(xì)胞反應(yīng)的 中央調(diào)節(jié)器 。
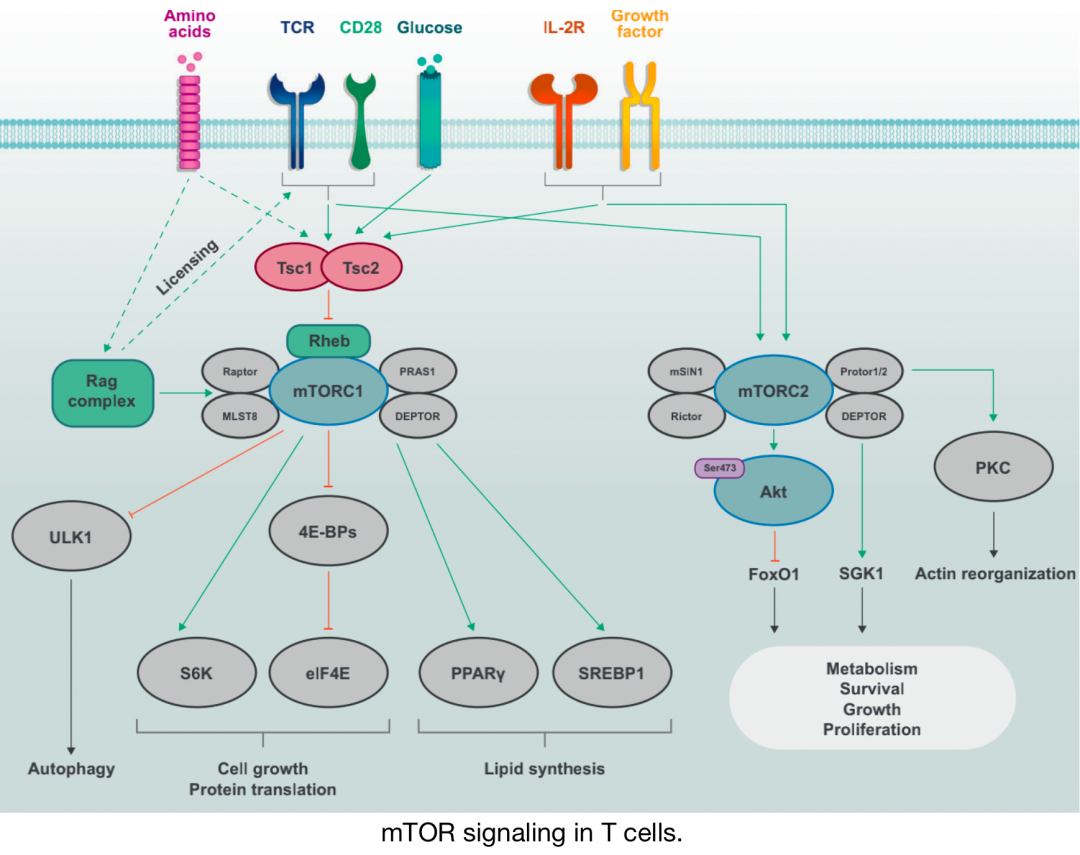
T細(xì)胞中的mTOR信號
離散的mTOR復(fù)合物mTORC1 (由mTOR、Raptor、PRAS1、DEPTOR和MLST8組成) 和mTORC2 (由mTOR、Rictor、Protor1/2、mSIN1和DEPTOR組成) 由免疫受體 (TCR、CD28和IL-2R) 和生長因子激活。mTORC1的激活也 對氨基酸等營養(yǎng)素敏感 ,氨基酸通過Rag復(fù)合物促進(jìn)mTORC1激活, Rag復(fù)合物 也起到允許TCR和CD28共刺激信號誘導(dǎo)mTORC1激活的“許可”作用。 Tsc復(fù)合物 的活性受到免疫和生長因子信號的抑制,通過抑制小G蛋白Rheb的激活,后者促進(jìn)mTORC1的激活。mTORC1通過S6K和eIF4E誘導(dǎo)細(xì)胞生長和蛋白質(zhì)翻譯,并通過PPARγ和SREBP1誘導(dǎo)脂質(zhì)合成。在營養(yǎng)充足的條件下,mTORC1通過ULK1抑制自噬。mTORC2主要在細(xì)胞生長、增殖和存活的環(huán)境或亞特異性代謝重編程中起著關(guān)鍵作用。
LKB1–AMPK信號通路 在調(diào)節(jié)細(xì)胞 代謝、增殖和生存以應(yīng)對營養(yǎng)和能量需求的改變方面起著核心作用 。LKB1–AMPK信號可促進(jìn)產(chǎn)生ATP的分解代謝途徑 ,并使T細(xì)胞在應(yīng)對能量應(yīng)激時具有代謝可塑性 。通過調(diào)節(jié)代謝重編程,LKB1和AMPK有助于T細(xì)胞的分化和功能 。在以下部分,我們將介紹LKB1和AMPK活性是如何調(diào)節(jié)的,其對代謝的影響以及在T細(xì)胞介導(dǎo)的免疫中的作用。
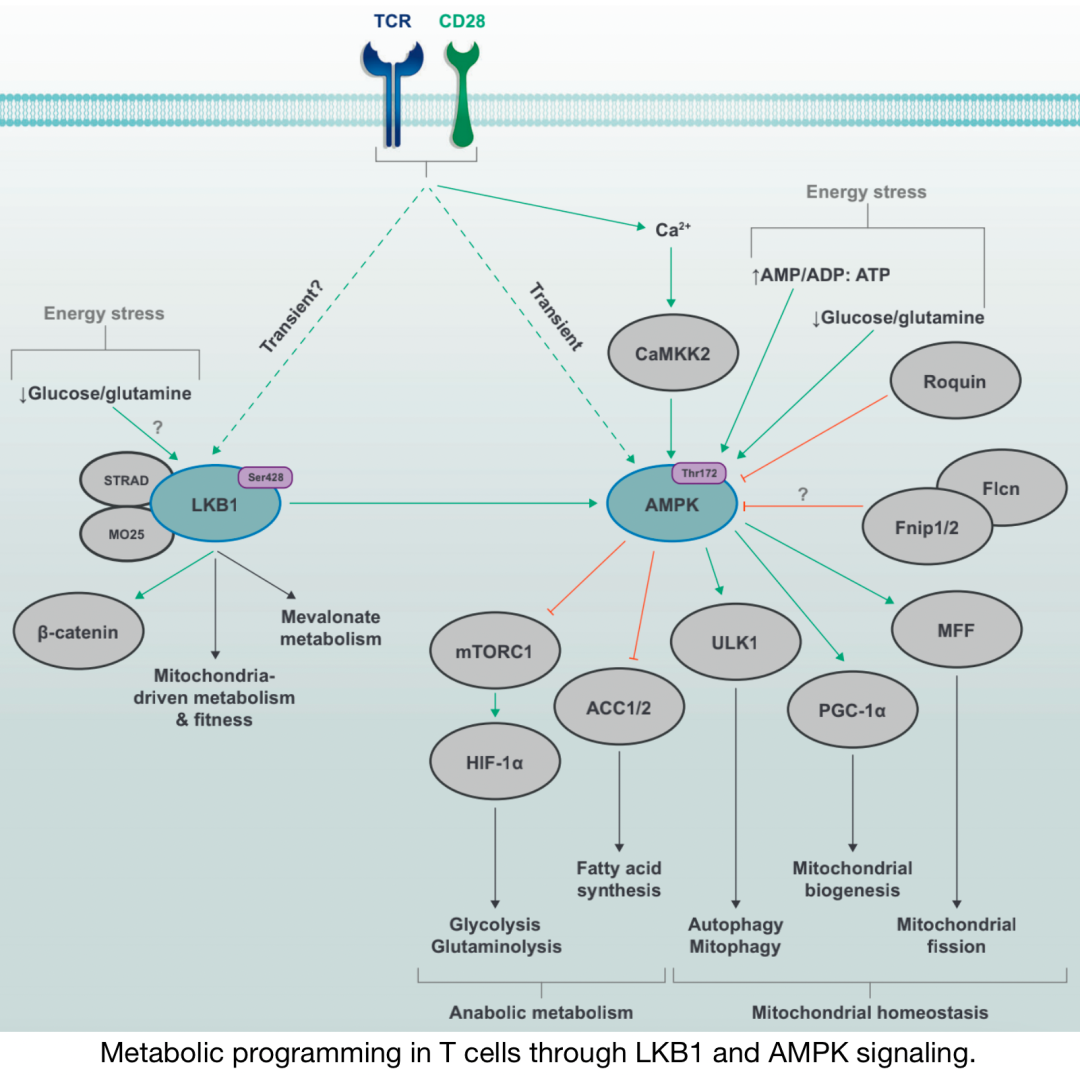
通過 L KB1和 AMPK 信號在 T 細(xì)胞中進(jìn)行代謝編程
能量應(yīng)激途徑激酶LKB1和AMPK由TCR和CD28共刺激信號激活,AMPK活性部分 由Ca2+CAMMK2途徑 介導(dǎo)。在缺乏葡萄糖或谷氨酰胺或AMP/ADP/ATP比值失衡 ,也可促進(jìn)LKB1–AMPK信號傳導(dǎo) 。上游營養(yǎng)感應(yīng)蛋白 ,如Fnip–Flcn復(fù)合物和Roquin,可以抑制AMPK功能 。LKB1的激活與線粒體代謝和適應(yīng)度的變化 以及特定環(huán)境下甲羥戊酸 代謝的增加有關(guān)。通過調(diào)節(jié)多個下游靶點(diǎn)的活性,AMPK信號可阻止糖酵解、谷氨酰胺解和脂肪酸合成的代謝程序,同時促進(jìn)分解代謝過程 ,如有絲分裂吞噬和自噬。AMPK還通過促進(jìn)線粒體凋亡 來驅(qū)動線粒體生物發(fā)生和線粒體動力學(xué),從而支持線粒體活性。
▉3.1? LKB1和AMPK的調(diào)節(jié)
《The LKB1-AMPK pathway: metabolism and growth control in tumour suppression》
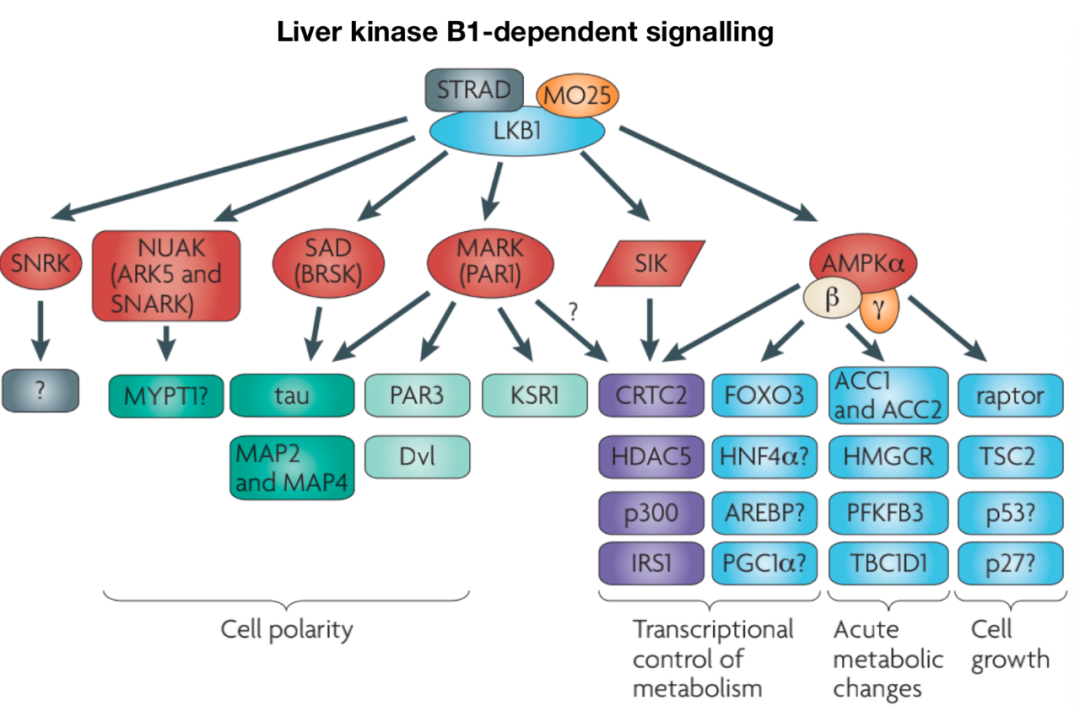
LKB1 是一種絲氨酸/蘇氨酸激酶,具有腫瘤抑制功能,參與調(diào)節(jié)細(xì)胞代謝和增殖。LKB1有許多下游靶點(diǎn) ,包括定義最明確的靶點(diǎn)AMPK和AMPK相關(guān)激酶 ,如BRSK、NUAK和MARK。LKB1的上游調(diào)節(jié)由細(xì)胞定位和翻譯后修飾介導(dǎo)。LKB1與STE20相關(guān)適配器 (STRAD) 和MO25形成復(fù)合物 ,分別促進(jìn)LKB1的細(xì)胞質(zhì)定位和激酶活性 。此外, Akt可以通過促進(jìn)LKB1核滯留來抑制LKB1 。
?
AMPK 是一種保守的絲氨酸/蘇氨酸激酶,由α、β和γ亞基組成。AMP和ADP與CBS3重復(fù)序列的結(jié)合限制了磷酸酶對AMPK催化α亞基上Thr172的訪問,該亞基是促進(jìn)AMPK激酶活性的關(guān)鍵磷酸化位點(diǎn)。最終,代謝產(chǎn)物的復(fù)雜調(diào)節(jié)允許AMPK活性 在不同能量應(yīng)激條件下的“分級”反應(yīng) 。至少三種上游激酶介導(dǎo)AMPK Thr172磷酸化:LKB1、鈣鈣調(diào)素激酶激酶2 (CaMKK2) 和TGF-β激活激酶1 (TAK1) ,其中LKB1和CaMKK2的作用最為清楚。在T細(xì)胞中,TCR和Ca2信號以及CD28共刺激協(xié)同作用,通過CaMKK2快速、瞬時激活A(yù)MPK。AMPK活性也通過蛋白質(zhì)-蛋白質(zhì)相互作用 進(jìn)行調(diào)節(jié)。Fnip1和Fnip2與AMPK形成復(fù)合物 ,F(xiàn)nip1的功能缺失突變提高了B細(xì)胞的AMPK活性,從而確立Fnip1是AMPK的負(fù)調(diào)節(jié)因子。Fnip結(jié)合蛋白Flcn也與AMPK相互作用,可能作為AMPK信號的負(fù)調(diào)節(jié)因子,有證據(jù)表明Fnip1和Flcn受AMPK的相互調(diào)節(jié)。
▉ 3.2 LKBI-AMPK介導(dǎo)的分解代謝
AMPK活性與 低營養(yǎng)豐度誘導(dǎo) 一致,通常與 分解代謝程序 有關(guān),以 生成ATP 。AMPK通過ULK1的磷酸化激活自噬,ULK1可促進(jìn)自噬介導(dǎo)的線粒體內(nèi)穩(wěn)態(tài),促進(jìn)T細(xì)胞存活。此外,AMPK獨(dú)立于mTOR調(diào)節(jié),在葡萄糖受限時,AMPK還通過Ser15處p53的磷酸化介導(dǎo)細(xì)胞周期阻滯。因此, LKB1–AMPK信號通過限制能量應(yīng)激期間的合成代謝和細(xì)胞生長促進(jìn)細(xì)胞存活 。此外,通過磷酸戊糖途徑產(chǎn)生的代謝物核酮-5-磷酸對LKB1–AMPK途徑的破壞促進(jìn)了脂肪生成。相反,LKB1也可以促進(jìn)甲羥戊酸代謝,以AMPK非依賴性方式支持Treg細(xì)胞內(nèi)穩(wěn)態(tài)。因此,LKB1–AMPK信號主要作用于 抑制脂質(zhì)合成途徑 ,但 AMPK依賴性LKB1信號可能在選擇性情況下促進(jìn)新生脂肪生成 。
?
對腫瘤細(xì)胞的研究表明, LKB1或AMPK信號的破壞促進(jìn)有氧糖酵解 ,部分是通過HIF-1α、因此增加糖酵解酶的轉(zhuǎn)錄。HIF-1α的LKB1–AMPK依賴性調(diào)節(jié)也可能部分依賴于mTORC1的抑制。LKB1–AMPK信號可能通過HIF-1α或ACC1介導(dǎo)的糖酵解和線粒體氧化代謝變化間接協(xié)調(diào)Th17和Treg細(xì)胞系的分化。此外,最近的研究表明,LKB1促進(jìn)穩(wěn)定的Foxp3表達(dá),以及Th2樣Treg細(xì)胞的發(fā)育,獨(dú)立于AMPK和mTORC1–HIF-1α信號,但依賴于β-連環(huán)蛋白信號。LKB1信號是TCR介導(dǎo)的Treg細(xì)胞激活后線粒體功能和線粒體依賴性代謝程序所必需的,包括FAO或嘌呤和嘧啶代謝。這些發(fā)現(xiàn) 強(qiáng)調(diào)LKB1和AMPK協(xié)調(diào)代謝重編程以調(diào)節(jié)T細(xì)胞分化和Treg細(xì)胞功能 。
▉4.? 免疫代謝信號網(wǎng)絡(luò)
上述主要信號通路在功能上并不排斥,相反,它們 相互交織,相互作用 ,共同達(dá)到適當(dāng)?shù)?strong>代謝調(diào)節(jié),以滿足細(xì)胞功能的特定環(huán)境需求。因此,許多這些調(diào)節(jié)輸入?yún)R聚在共同的節(jié)點(diǎn) 上,包括單個分子和整個細(xì)胞過程 。
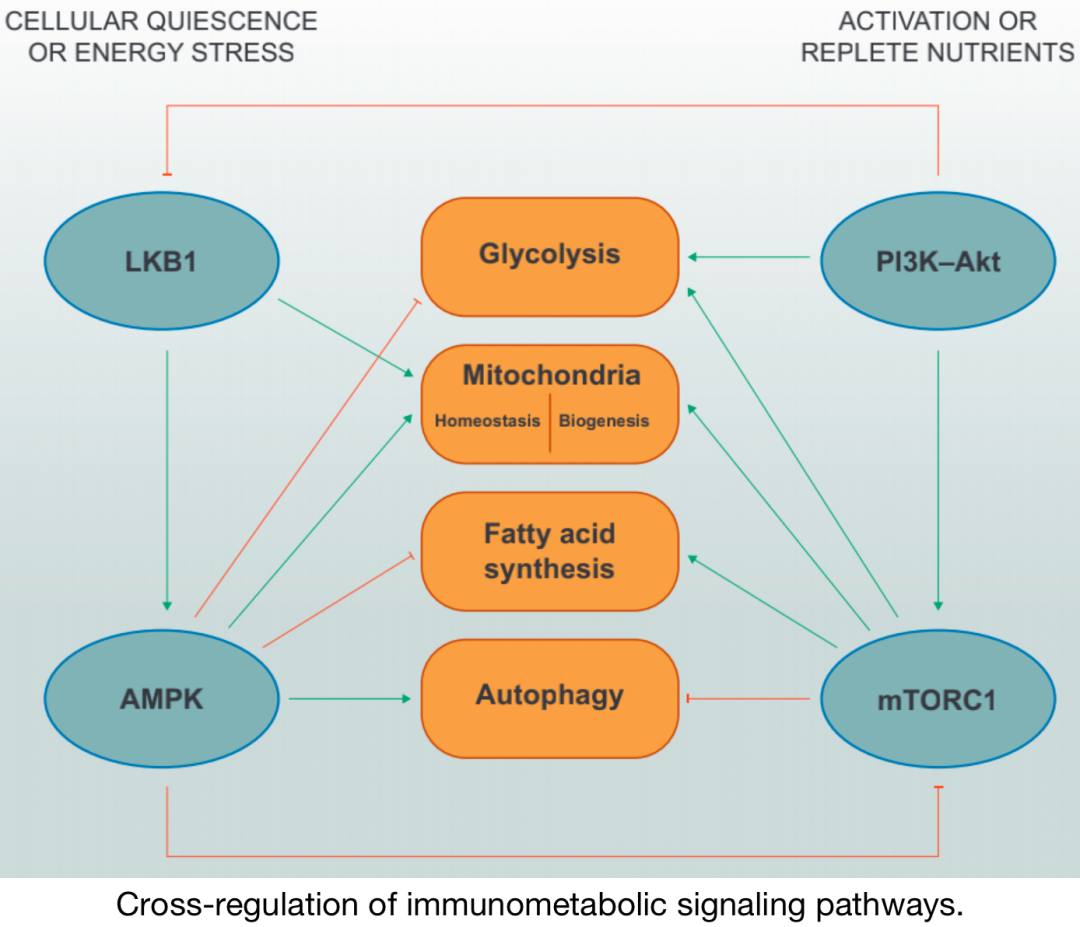
免疫代謝通路的交叉調(diào)節(jié)
在營養(yǎng)缺乏 的條件下,LKB1–AMPK 信號 抑制合成 代謝相關(guān)程序,如糖酵解和脂肪酸合成,同時促進(jìn)線粒體內(nèi)穩(wěn)態(tài)和自噬。AMPK通過其專一性轉(zhuǎn)接器蛋白Raptor的磷酸化直接抑制mTORC1。在激活和/或營養(yǎng)充足 的條件下,PI3K–Akt和mTORC1 信號促進(jìn) 糖酵解、線粒體生物發(fā)生和脂肪酸合成 ,同時抑制自噬 。據(jù)報道,Akt可以磷酸化LKB1 以抑制其功能定位 。
一般來說,PI3K–AGC激酶和mTOR活性的升高,以及LKB1–AMPK活性的降低 ,與細(xì)胞生長、增殖和效應(yīng)器功能 相關(guān),這與代謝程序的變化相關(guān)。 上游復(fù)合體的信號整合最終與下游代謝程序的調(diào)節(jié)有關(guān) ,決定免疫細(xì)胞代謝程序的信號通路也 受到代謝物和營養(yǎng)物質(zhì)的交互影響 。這種 “雙向代謝信號” 為 激酶依賴性信號和基因轉(zhuǎn)錄 提供了另一層調(diào)控,以控制不同環(huán)境下的免疫細(xì)胞功能 。
免疫信號網(wǎng)絡(luò) 在免疫代謝中的作用是一個令人興奮的研究領(lǐng)域,對治療和人類健康具有廣泛的意義。在這篇綜述中,我們強(qiáng)調(diào)了 PI3K–AGC、mTOR和LKB1–AMPK信號在T細(xì)胞功能和命運(yùn)中的關(guān)鍵作用 。該領(lǐng)域在理解 這些信號分子是如何調(diào)節(jié)的以及它們是如何重新編程T細(xì)胞代謝 方面取得了很大進(jìn)展,確定相對知名的 上游免疫信號 (如TCR、細(xì)胞因子受體) 如何在代謝中傳遞不同的功能程序 ,可能為改善過繼性T細(xì)胞治療 (如抗腫瘤CAR-T細(xì)胞治療) 提供新的 治療靶點(diǎn) 。
Saravia, J., Raynor, J.L., Chapman, N.M.?et al.?Signaling networks in immunometabolism.?Cell Res?30,?328–342 (2020). https://doi.org/10.1038/s41422-020-0301-1
編輯:小果果,轉(zhuǎn)載請注明出處:http://www.448371.com/linchuang/lcyj/19859.html
免責(zé)聲明:本站所轉(zhuǎn)載文章來源于其他平臺,主要目的在于分享行業(yè)相關(guān)知識,傳遞當(dāng)前最新資訊。圖片、文章版權(quán)均屬于原作者所有,如有侵權(quán),請及時告知,我們會在24小時內(nèi)刪除相關(guān)信息。
說明:本站所發(fā)布的案例均摘錄于文獻(xiàn),僅用于科普干細(xì)胞與再生醫(yī)學(xué)相關(guān)知識,不作為醫(yī)療建議。



 微信掃一掃
微信掃一掃  支付寶掃一掃
支付寶掃一掃 