六.來自ICB和CAR-T細胞的免疫治療生物標志物
癌癥是高度復雜的疾病,其特點是不僅有惡性細胞的過度生長,而且有免疫反應的改變。免疫系統的抑制和重新編程在腫瘤的發生和發展中起著關鍵作用, 免疫治療的目的是重新激活抗腫瘤免疫細胞,克服腫瘤的免疫逃逸機制。 有證據表明,多種免疫治療方法的結合可能是提高治療效果的途徑之一。
然而,腫瘤免疫治療 今天分享一篇發表在Signal Transduction and Targeted Therapy(IF=38.104)的綜述文章。這篇綜述描述了 免疫系統對抗癌癥的機制,并討論了可以促進抗腫瘤免疫反應的治療靶點和細胞類型。
1974年,白細胞介素(IL)-2 被發現在T細胞分化和生長中起著至關重要的作用。T細胞的激活是抗病毒和抗腫瘤適應性免疫中的一個關鍵事件,它主要是通過雙信號途徑完成的。第一個信號是抗原特異性信號,包括T細胞表面受體(TCR)與抗原肽-主要組織細胞復合體(MHC)的特異性結合。第二個信號是由T細胞與抗原提呈細胞(APC)表面的刺激分子(CMs)的交流介導的。 在確定了負責抗原識別的T細胞受體(TCRs)后,1986年,科學家們發現了在活化T細胞上表達的分子CD28。隨后,人們發現T細胞的激活需要來自TCR和CD28的信號,此后CD28被命名為?“共刺激分子”。Pierre Golstain的團隊發現了一種與CD28結構相似的蛋白質;它被命名為細胞毒性T淋巴細胞相關抗原4(CTLA-4),并被假設為一種潛在的T細胞激活分子。 1994年底,Allison團隊報告了阻斷CTLA-4可以增加T細胞的抗腫瘤活性和抑制腫瘤生長的突破性發現。因此,第一次證明了抑制一種負性免疫調節劑可以抑制腫瘤的發展;這種方法后來被Allison命名為?“免疫檢查點阻斷”(ICB)。2011年,第一個針對CTLA-4的抗體ipilimumab被批準用于黑色素瘤治療,成為第一個免疫檢查點(IC)抑制劑。 20多年前,京都大學Tasuku Honjo的研究小組發現了程序性細胞死亡蛋白1(PD-1)。PD-1基因敲除導致小鼠出現自身免疫性疾病和異常激活的免疫細胞,表明其具有免疫抑制作用。 2000年,B7-H1被確定為PD-1的配體,因此獲得了它的第二個名字PD-L1。這些發現表明,PD-1是另一種IC。文章總結了癌癥免疫療法在一段時間內的主要里程碑式的突破(圖1)。針對CTLA-4和PD-1的抗體以及CAR-T細胞療法代表了有希望的方法,在這些方法中,免疫系統的某些成分可以被操縱,以扭轉抑制并針對腫瘤。 然而并非所有患者都對這些療法有反應,這表明腫瘤引起的免疫改變的復雜性。Robert Schreiber博士提出了?“癌癥免疫編輯?“ 的概念,描述了惡性細胞如何對最初的免疫識別做出反應,隨后發展出逃避機制,甚至 “重塑 “免疫系統,成為致癌物質。這樣的免疫編輯過程幾乎在體內或全身免疫細胞存在時都會發生,從而形成高度抑制的腫瘤微環境(TME)。尋找激活抗腫瘤T細胞反應和針對抑制性TME的協同方法一直是腫瘤免疫治療的主要研究重點。
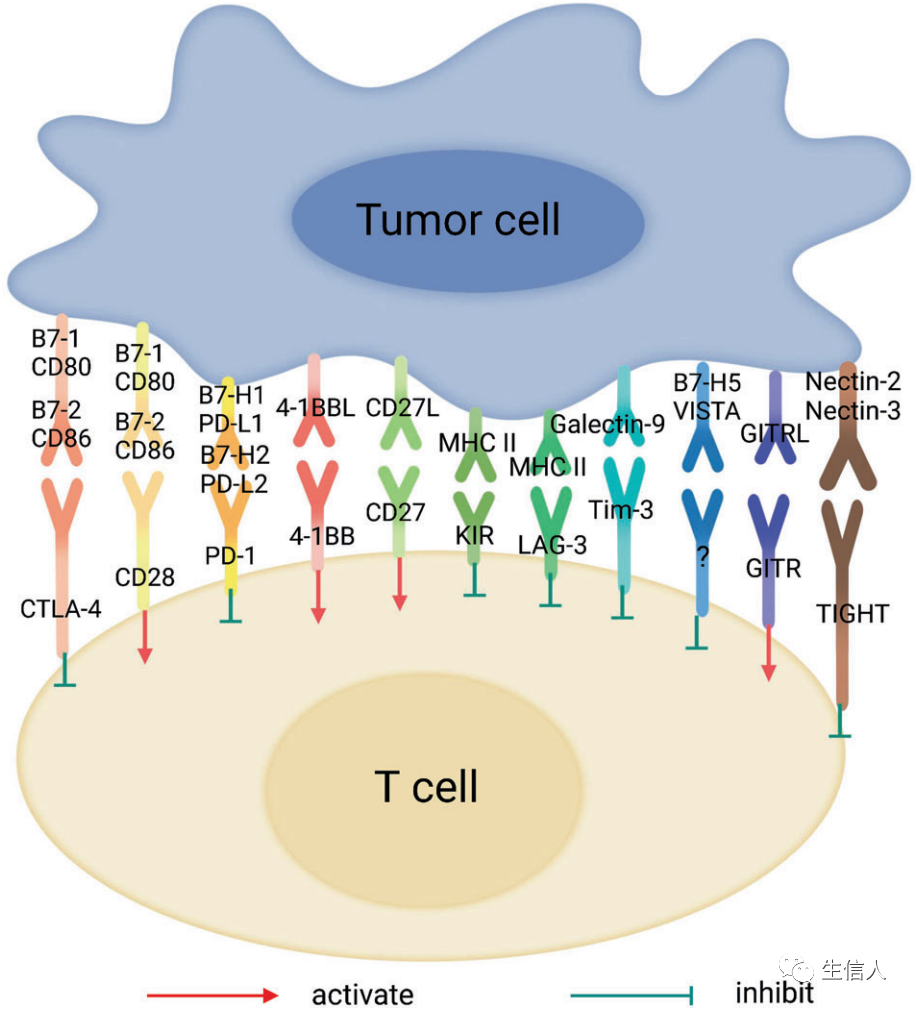
ICs是一類免疫抑制分子,在免疫細胞上表達,可以抑制免疫細胞的激活,因此在預防自身免疫中起著關鍵作用 (圖2)。相反,ICs的過度表達會抑制免疫功能,促進腫瘤的發生。因此,ICB療法通過阻斷ICs和增強抗腫瘤T細胞活性來抑制腫瘤生長。ICB的發展是通過靶向PD-1/PD-L1和CTLA-4/B7-1/2這兩條IC通路開始的,對它們的阻斷已經取得了顯著的臨床進展。 迄今為止,T細胞上其他幾個通過不同機制介導抑制性信號的IC分子已被發現,并有可能被作為癌癥免疫治療的靶點加以利用。
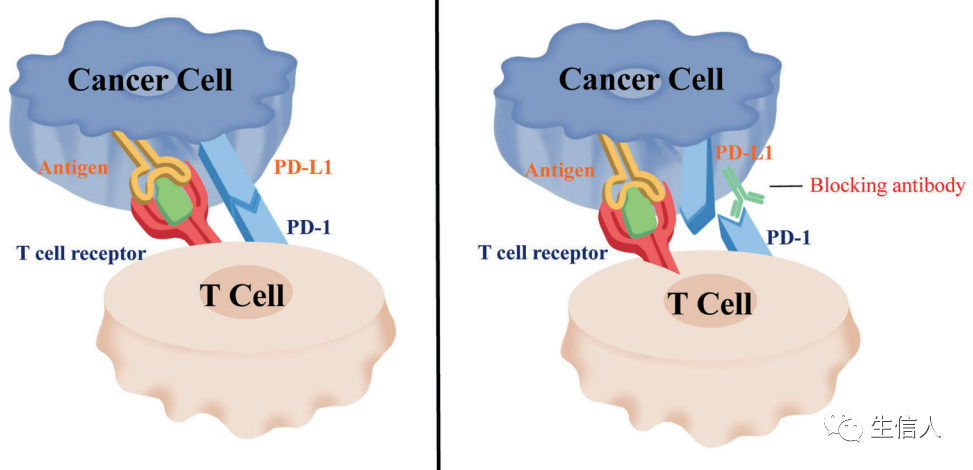
研究發現PD-1能抑制T淋巴細胞的功能,這對控制自身免疫反應至關重要。PD-L1(最初被確定為B7-H1),在多種類型的腫瘤上高度表達,可與PD-1結合并介導腫瘤的免疫逃逸。因此,對PD-1的抑制可以重新激活T細胞功能。PD-1不僅在T細胞上表達,而且還在NK細胞、B淋巴細胞、巨噬細胞和樹突狀細胞(DCs)上表達,表明PD-1可能在重塑腫瘤免疫微環境甚至全身性抗腫瘤免疫中發揮非常有效的作用。 PD-1抑制劑可以特異性地與PD-1結合,從而減弱對T淋巴細胞的免疫抑制調節,使T淋巴細胞能夠參與殺傷腫瘤細胞(圖3)。 ?
CTLA-4也被稱為CD152,是一種跨膜蛋白,在活化的CD4+和CD8+T細胞中表達。CTLA-4和CD80/CD86的結合可以抑制T細胞的激活信號,防止自身免疫性疾病。阻斷CTLA-4可以直接針對效應性T細胞的抑制信號,減少Tregs的抑制作用,從而有效提高T細胞的抗腫瘤作用。 盡管兩者都是有代表性的IC分子,CTLA-4和PD-1以不同的方式調節T細胞功能(圖4)。CTLA-4的抑制性信號消極地調節T細胞的啟動,而PD-1主要介導啟動后的T細胞的激活和增殖。在腫瘤方面,人們發現針對PD-1的ICB通常會導致現有抗腫瘤T細胞的擴增和招募,而抗CTLA-4治療會產生新的T細胞克隆。這些結果表明,CTLA-4和PD-1可以同時作為靶點,發揮協同抗腫瘤作用。
圖4. CTLA-4和PD-1的圖解
T細胞免疫球蛋白域和粘蛋白域-3(Tim-3,CD366)是一種T細胞表面抑制性分子,主要表達在CD4 + T輔助細胞1(Th1)和CD8 + CTL細胞以及具有強化抑制功能的Treg細胞亞群上。 后來發現Tim-3,也被稱為HAVCR2,也表達在一些先天性免疫細胞上,包括樹突狀細胞、NK細胞、單核細胞和巨噬細胞。 在抗原刺激下,LAG-3可以在CD4 +和CD8 + T細胞上被誘導。長期感染病毒、細菌和寄生蟲導致持續暴露于抗原,這導致LAG-3的高水平和持續表達,隨后細胞因子釋放、細胞溶解活性和增殖潛力減少。在一些小鼠腫瘤模型中觀察到LAG-3和PD-1在瘤內T細胞上的共同表達,當結合這兩種分子的阻斷抗體時,觀察到對腫瘤生長的協同抑制作用。 LAG-3因此成為癌癥免疫治療最關鍵的新靶點之一。Relatlimab是第一個進入臨床的LAG-3的抑制劑,它能阻斷LAG-3與MHC II的相互作用。纖維蛋白原樣蛋白1(FGL1)是Lag-3的配體。發現它與Lag-3結合形成一個新的PD-1/PD-L1獨立的免疫檢查點途徑,導致T細胞衰竭、功能障礙,以及腫瘤細胞逃避免疫監視。在抗PD-L1的基礎上阻斷FGL1,有可能成為臨床實踐中的另一種新型ICB策略,尤其是在非小細胞肺癌(NSCLC)的靶向治療方面。 核受體亞家族2群F成員6 (NR2F6)最近被報道為細胞內IC分子,是淋巴細胞固有的核受體。NR2F6作為轉錄因子調節與腫瘤抗原特異性T細胞反應相關的細胞的激活、募集、增殖和內穩態。在非小細胞肺癌組織的腫瘤浸潤淋巴細胞(TILs)中檢測到NR2F6的高表達 ,NR2F6表達上調與細胞因子(IL-2、TNF-α和IFN-γ)的產生受損相關,提示TILs上的NR2F6有助于腫瘤免疫抑制。此外,NR2F6的破壞導致腫瘤抑制,并增強PD-L1阻斷劑在腫瘤治療中的作用,提示NR2F6抑制劑可能成為一種新的免疫療法,可以克服對現有ICB治療的耐藥性。 T細胞免疫球蛋白和ITIM域蛋白(TIGIT)是一種I型跨膜蛋白。TIGIT屬于免疫球蛋白超家族(IgSF),可以在T細胞、調節性T細胞、記憶性T細胞和NK細胞上表達。TIGIT通過與表達在抗原呈遞細胞(APC)上的配體CD155和CD112相互作用,介導對NK細胞和T細胞激活的抑制作用。在人類腫瘤中,TIGIT被發現與多個IC分子共表達,包括PD-1、TIM-3和LAG-3。 TIGIT、TIM-3和PD-1的共同表達與患者的生存率低有關。同時阻斷TIGIT和PD-1信號通路可以增加腫瘤特異性CD8+T細胞中IFN-γ和TNF-α的表達,支持抗TIGIT治療的發展。 V-集免疫調節受體(VISTA),也被稱為PD-1H或DD1α,是近年來發現的一種免疫調節蛋白。它主要表達于淋巴器官和骨髓細胞,其結構與PD-L1相似。研究表明, 表達VISTA的APC對CD4 +和CD8 + T細胞有抑制作用; 當這種分子被阻斷時,由T細胞介導的免疫功能得到了挽救,說明VISTA是一種抑制T細胞反應的IC分子 。在T細胞中,VISTA和PD-1的抑制作用是相互獨立的,在腫瘤小鼠模型中的研究也驗證了同時應用抗PD-1和抗VISTA抗體可以抑制腫瘤生長,延長生存期。 B和T淋巴細胞衰減劑(BTLA)屬于免疫球蛋白超家族。它在T細胞、靜止的B細胞、巨噬細胞、樹突狀細胞和NK細胞上表達,其結構和功能與PD-1和CTLA-4相似。BTLA的配體是皰疹病毒進入介質(HVEM)。 當BTLA與HVEM結合時,它產生抑制性信號并抑制T細胞的激活。抗BTLA治療可促進T細胞增殖,BTLA基因敲除的小鼠顯示出更高的免疫活性。
OX40,也被稱為CD134,是腫瘤壞死因子受體(TNFR)超家族的成員,在T細胞激活后24-72小時表達。它的配體OX40L,也被稱為CD252,主要表達在活化的APC表面。OX40-OX40L的相互作用可以啟動T細胞激活信號以及細胞周期蛋白A、Bcl-2抗凋亡分子、細胞因子和細胞因子受體的表達。小鼠模型顯示,刺激OX40的特異性抗體可以減少Tregs的數量,從而保持效應T細胞的功能,并顯示出高抗腫瘤活性有許多針對OX40-OX40L途徑的臨床研究,包括對激發OX40的特異性抗體的單藥應用或與化療、放療、手術、小分子靶向治療、細胞因子或其他ICB藥物的組合。 誘導共刺激分子(ICOS),又稱CD278,是免疫球蛋白超家族的一員。ICOS在活化的T細胞表面表達,調節T細胞增殖和功能。ICOS的激活依賴于其配體ICOS- l,主要在B細胞和APCs中表達。ICOS已被證明是ICB療效的一個重要標志。當anti-CLTA-4治療惡性黑色素瘤時,發現ICOS + CD4 + T細胞的豐度與較好的療效相關。在小鼠模型中,單獨使用ICOS激動劑很難引起足夠的抗黑色素瘤反應;但ICOS與anti-CLTA-4之間存在協同效應; 此外,ICOS敲除小鼠對anti-CLTA-4治療反應較差。 4-1BB,也被稱為CD137,是TNFR家族的一員。4-1BB在調節性T細胞上的作用是復雜的,研究結果相互矛盾。但重要的是,4-1BB基因敲除小鼠發生自身免疫性疾病,提示其在免疫平衡中發揮重要作用,有可能被靶向引發腫瘤特異性免疫識別。 與TNFR家族的其他成員不同,CD27只在淋巴細胞表面表達,包括新生和活化的CD4+和CD8+T細胞。 當它與其配體CD70相互作用時,CD27誘導效應性和記憶性T細胞的增殖和分化,并增強B細胞和NK細胞的激活。小鼠模型表明,誘導CD27信號通路可以抑制腫瘤的生長。CD27/CD70信號通路的抗腫瘤免疫作用。Varlilumab是一種針對CD27的人源化單克隆抗體,可促進細胞因子的產生和T細胞的激活。在I期臨床研究中,Varlilumab在晚期實體瘤患者中的耐受性良好,并顯示出初步的安全性結果。除了單藥治療外,varlilumab還與抗PD-L1抗體聯合使用。
在人體內,NK細胞主要以CD3- CD56 +淋巴細胞群為特征,而CD16 + CD56dim亞型主要存在于血液中。NK細胞作為自然免疫系統的重要組成部分,在去除衰老細胞和致病微生物方面發揮著重要作用。 NK細胞不像TCRs那樣通過特定的受體識別靶細胞;它們通過種系基因表達的受體識別細胞。NK細胞的負調控因子包括KIRs(免疫球蛋白樣受體)、CD94-NKG2和MHC-I。 在腫瘤免疫學的背景下,腫瘤細胞通過下調MHC的表達來逃避獲得性免疫,從而更容易受到NK細胞的細胞毒性作用。此外,在抗體治療中,NK細胞在介導抗體誘導的細胞毒性(ADCC)中發揮著重要作用。NK細胞還可以通過分泌細胞因子或招募樹突狀細胞、巨噬細胞和T細胞等免疫細胞參與清除腫瘤細胞的過程,直接發揮抗腫瘤作用238,使其成為癌癥免疫治療的誘人靶點。 殺傷細胞免疫球蛋白樣受體(KIR)家族是一類高度多態性的分子,主要表達在一些NK細胞和T細胞的表面,可分為多個亞型。其中,KIR2DTl1-3和KIR3DL1通過結合MHC分子(HLA-C/HLAB)發揮抑制作用。由于基因多態性高的特點, 多個KIR基因及其配體的結合可引起多種疾病,包括自身免疫性疾病,特別是部分KIR基因與特異性配體的結合,可增加癌癥的風險。 在小鼠模型中,靶向活化的NK細胞表面受體KIR2DS2的治療顯示出明顯優于靶向傳統共刺激分子的抗腫瘤活性。 NK細胞凝集素樣受體亞家族C成員1 (NKG2A)是NKG2家族的“抑制性”成員,主要在CD56 hi NK細胞、NKT細胞和CD8 + αβ t細胞亞群中表達。它與CD94形成異二聚體受體,并與它的配體結合,非經典的MHC I分子HLA-E,在大多數正常組織中表達。NKG2A/CD94與HLA-E的相互作用可抑制NK細胞和T細胞的激活,表明其作為IC分子的靶向性。 Monalizumab (IPH2201)是一種NKG2A單克隆拮抗抗體,可阻斷NKG2A與HLA-E之間的相互作用,已在白血病小鼠模型中顯示治療效果。 CD96是NK細胞上表達的免疫球蛋白超家族的一員,可識別配體CD155。CD96在腫瘤浸潤NK細胞上的表達高于周圍組織NK細胞。肝細胞癌樣本中NK細胞CD96表達水平較高預示預后較差。
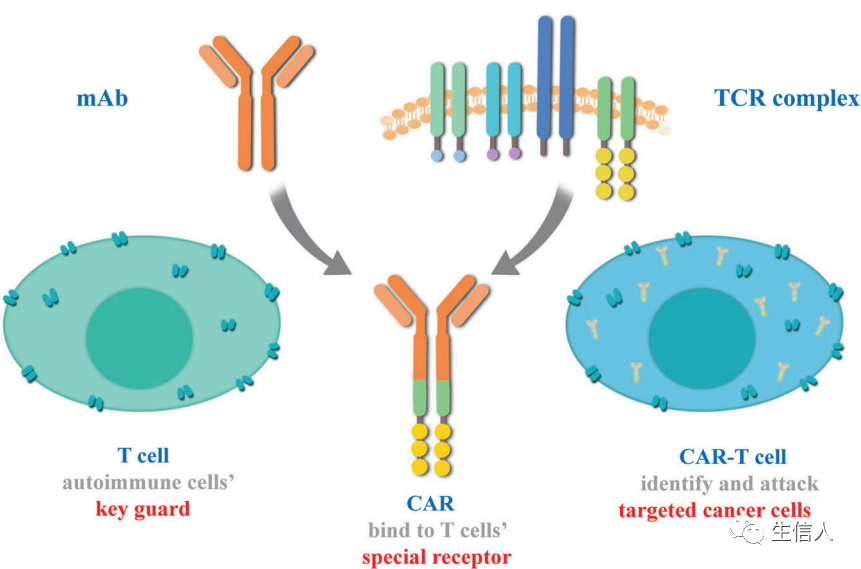
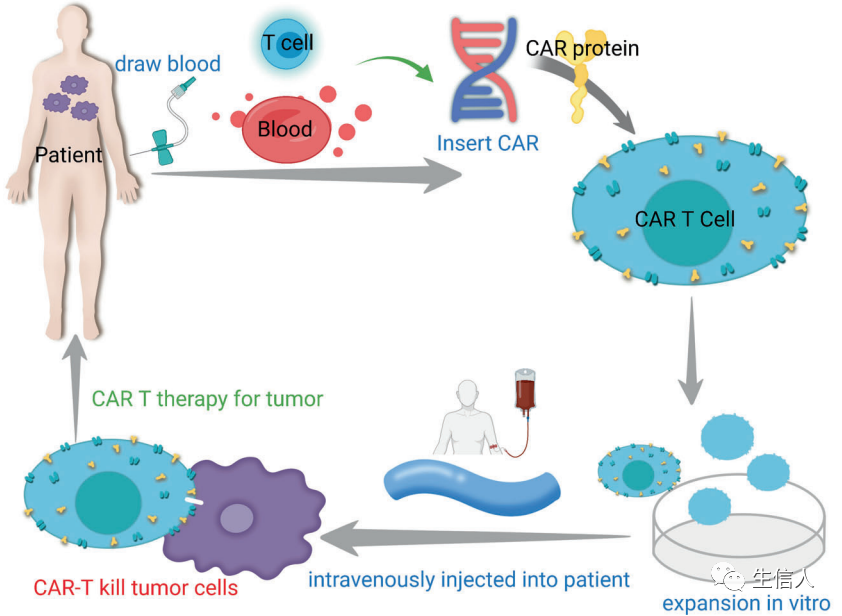
具有細胞毒性潛能的免疫細胞,包括T細胞、NK細胞和巨噬細胞,能夠識別并清除感染或受損的細胞。T細胞的細胞毒性作用不同于其他細胞,因為它具有抗原特異性。細胞免疫療法,也稱為過繼細胞轉移(adoptive cell transfer,ACT),利用這些類型的免疫細胞的殺傷能力來治療癌癥。在此,作者討論已經取得重大研究和臨床進展的四種主要ACT類型: CAR – T細胞治療、TIL治療、工程TCR治療和NK細胞治療。 TILs是可以從腫瘤組織中識別和純化的異質性淋巴細胞,它們的豐富程度已被發現與較好的預后相關。TILs是第一批被用于ACT的細胞之一。這些細胞可以從腫瘤中分離出來,在實驗室環境中進行體外擴增,并大量重新注射到癌癥患者體內以消除腫瘤細胞。TIL療法已經在臨床研究中進行了嚴格的測試,作者總結了TIL療法針對不同類型腫瘤的最新進展,包括:黑素瘤、肺癌、宮頸癌、轉移性乳腺癌、骨肉瘤、卵巢癌。 TCRs是T細胞表面的特異性受體。通過識別和結合MHC呈遞的抗原,激活T細胞的分裂和分化。然而,并不是所有病人都有能識別腫瘤的T細胞。因此,TCR-T療法從患者身上提取T細胞,并將這些細胞擴大,使患者擁有能夠識別特定癌癥抗原的新的TCR。用于TCR-T治療的工程TCRs的設計高度依賴于特異性腫瘤抗原的識別。一些抗原,如NY-ESO-1,在腫瘤組織中廣泛表達,可用于開發TCRs治療不同類型的腫瘤。然而,TCR可以以患者特異性的方式進行識別和合成。TIL治療利用了瘤內淋巴細胞與全身淋巴細胞之間的差異,對患者腫瘤中特定突變的識別指導了TCRs的生成和應用,從而有效地靶向這些突變。然后,這些TCRs可以被分離、克隆并在T細胞上表達,然后這些工程T細胞在體外擴增并重新注入患者體內。這是一種高度個性化的治療方法,增強了治療的特異性。TCR療法在黑色素瘤的治療上取得了突破性進展,在肝癌、乳腺癌、卵巢癌的治療上也取得了一定成效。然而,TCR對腫瘤抗原的識別需要MHC分子的抗原表達,腫瘤細胞會通過降低MHC的表達來逃避T細胞的殺傷。 CAR-T細胞治療是另一種ACT策略。與TCR T療法的原理相似,患者的T細胞“配備”了合成的CAR,擴展并重新注入患者體內,以產生腫瘤特異性免疫反應(圖5)。CAR被設計用于識別腫瘤相關抗原(TAAs),其獨立于MHC呈遞,因此使T細胞能夠以MHC不受限制的方式識別癌細胞。
CAR-T細胞是通過在T細胞的質膜上表達腫瘤特異性CARs而產生的。? CARs的結構通常包括三個部分:細胞外抗原結合域、連接體/跨膜域、和細胞內信號傳導域。 細胞外抗原結合區是利用抗體、配體和肽的序列設計的,以便與TAAs特異性結合。跨膜結構域負責連接細胞外結合結構域和細胞內信號結構域并將其固定在細胞膜上。細胞內信號區,包括CD3-zeta結構域和共刺激結構域,將抗原識別的信號轉發給細胞,介導T細胞的激活。CAR-T細胞治療涉及合成生物學(CAR設計)、病毒技術(CAR轉導)和細胞制造(CAR-T細胞擴增)的整合(圖6)。CAR的引入在工程T細胞中產生了腫瘤特異性激活潛力,而體外培養和擴增允許繞過腫瘤引起的免疫抑制。因此,大量的腫瘤特異性細胞被輸回病人體內。CAR-T細胞療法對幾種類型的癌癥顯示出有希望的臨床結果。
圖6. CAR-T治療的工作流程
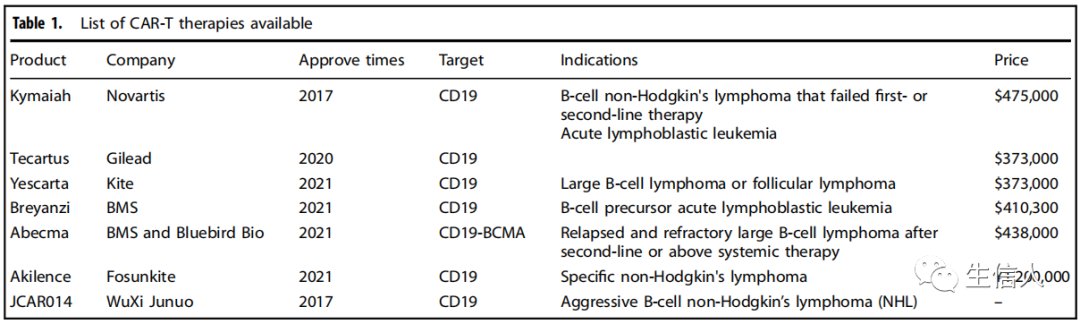
CAR-T細胞治療的更廣泛應用仍然面臨著來自不同方面的挑戰。首先,當CAR-T細胞攻擊腫瘤細胞時,可能會引起嚴重的副作用和毒性,可能是致命的。其次,一些CAR-T細胞的細胞毒性不是高度腫瘤特異性的,可能會對正常組織造成損傷。第三,大多數CAR-T細胞產品的制造過程是耗時的,這可能會導致一些患者的腫瘤在細胞生產窗口期進一步惡化。此外,CAR-T治療血癌的長期療效仍需長期隨訪觀察,CAR-T細胞治療實體瘤的應用有待進一步研究。這些挑戰將決定未來整個T細胞工程領域的發展。 細胞因子釋放綜合征(CRS),也被稱為“細胞因子風暴”,是CAR-T治療最常見的不良反應。 大多數CAR-T細胞的臨床研究表明,長期療效不確定,因為癌細胞表達的各種免疫逃逸因子導致T細胞衰老和衰竭。針對實體瘤的CAR-T細胞面臨的挑戰包括阻斷T細胞浸潤的血管疾病,有限的TAA選擇和腫瘤異質性,這導致了抗原逃逸。CAR-T細胞在實體瘤中的應用需要進一步探索。 利用特異性和廣泛的靶向性: CAR-T細胞的長期抗腫瘤功能因輸注后腫瘤復發而復雜化。考慮到CAR-T治療后抗原逃逸的風險,設計針對多個TAAs的雙特異性CAR-T是可以采用的。提高CAR-T細胞的適應度,包括活化潛能、增殖能力和生存能力,延長CAR-T細胞在患者體內的生存時間,是提高臨床療效的關鍵方向之一。 降低制造成本: 到目前為止,所有批準的CAR-T細胞療法都使用自體T細胞來產生治療產品。生產這種高度個性化的療法需要很高的成本。除了高成本外,CAR-T細胞療法的質量和穩定性一直是人們關注的主要問題。自體T細胞在質量和數量上不一致,特別是在接受過大量放療和化療的患者中。 毒性控制: CAR-T細胞治療顯示的毒性和副作用表明需要制定一些控制程序來調節CARs的活性。大量的方法已經被用來控制CAR-T細胞的安全性; 其中包括通過安裝自殺開關快速清除注入的細胞,這種開關可以由小分子或抗體控制。常用的自殺開關包括誘導型caspase-9 (iCasp9)、單純皰疹病毒中的胸苷激酶(HSV-TK)和自殺表位。然而,這樣的自殺開關清除了所有的治療CAR-T細胞,從而降低了抗腫瘤反應。因此,不清除CAR-T細胞的非細胞毒性可逆系統正在開發中,并具有保持細胞毒性和控制毒性反應之間平衡的潛力?(表1) 。
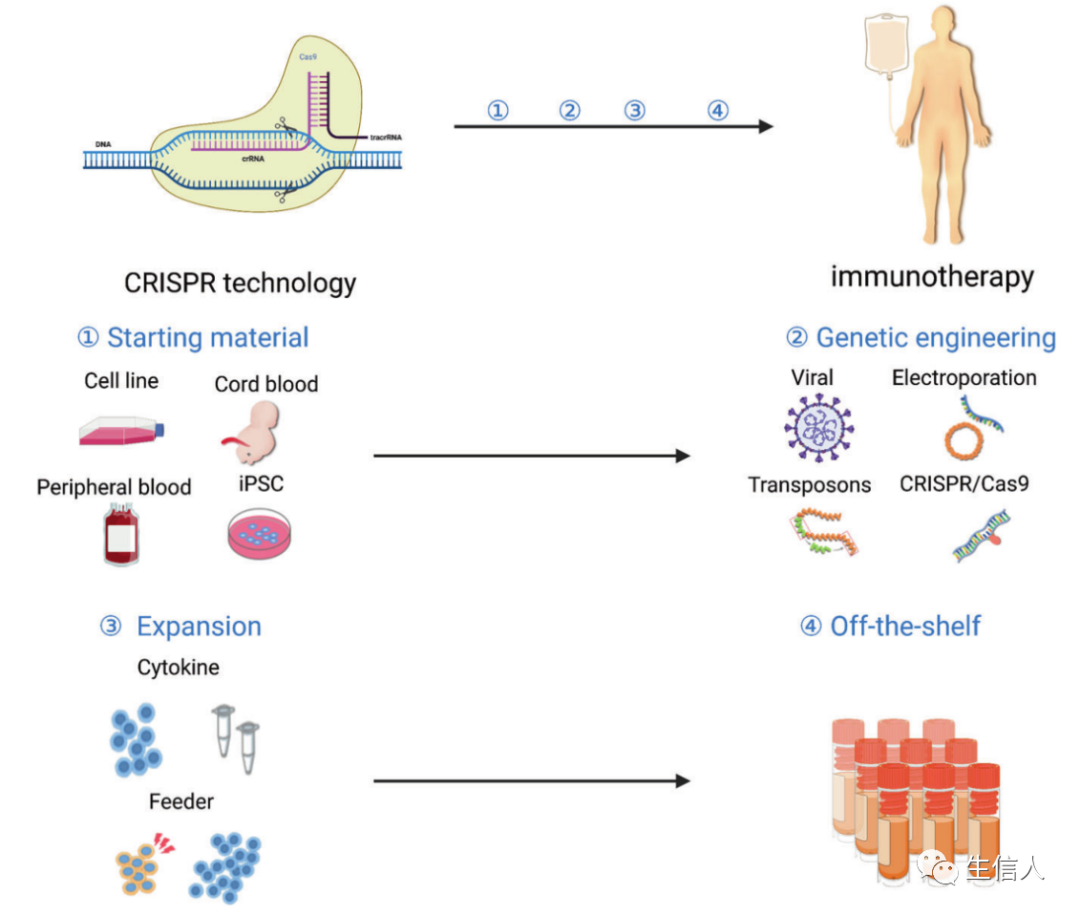
NK細胞是另一種重要的免疫細胞類型,可以介導直接細胞毒性。從機制上講,NK細胞在對抗癌癥的第一道防線上發揮著關鍵作用,通過兩種途徑介導抗腫瘤作用: 通過釋放穿孔后蛋白和顆粒酶或死亡受體的直接細胞毒性,以及通過分泌激活APCs和T細胞的細胞因子和趨化因子的調節作用。因此,除了本文前面討論的以NK細胞上的ICs為靶點的藥物外,利用NK細胞的ACT也在快速發展中。 與T細胞類似,NK細胞也可以轉導表達CARs。CAR-NK細胞的發展緊隨CAR-T細胞治療的演變,CAR-NK細胞往往直接采用CAR-T細胞的設計。 許多臨床前研究已經證實了CAR-NK細胞針對其他類型腫瘤的抗腫瘤活性。由于NK的細胞毒性是由 “缺失自我 “的識別引發的,因此NK細胞尤其具有殺死MHC下調的腫瘤細胞的能力。NK細胞對病毒感染的細胞也有特殊的殺傷能力,因此特別適合于治療HPV或EBV相關的腫瘤。 NK細胞毒性可受多種免疫抑制機制的影響,包括IL-10,吲哚胺2,3-雙加氧酶,前列腺素E2,轉化生長因子β (TGF-β)和缺氧。增強NK細胞的細胞毒性和在體內的持久性被認為是推進NK治療的主要方向。 支持NK細胞維持的細胞因子:IL-15已被確定為增強NK細胞活性的關鍵細胞因子。 在黑色素瘤、結直腸癌、淋巴瘤和肺癌等癌癥的同基因小鼠模型中,IL-15的注射具有良好的耐受性,并促進NK細胞的擴增。隨著對NK細胞活化和維持的研究越來越深入,未來的治療方法不僅必須產生腫瘤特異性NK細胞,還必須增加其在體內的持久性,以增強其治療效力。 近年來,CRISPR/Cas9技術極大地提高了我們對腫瘤基因組學的認識,并為癌癥免疫治療做出了貢獻。使用該基因組編輯系統,可進一步改造治療性免疫細胞,以增強腫瘤識別并減少衰竭(圖7)。
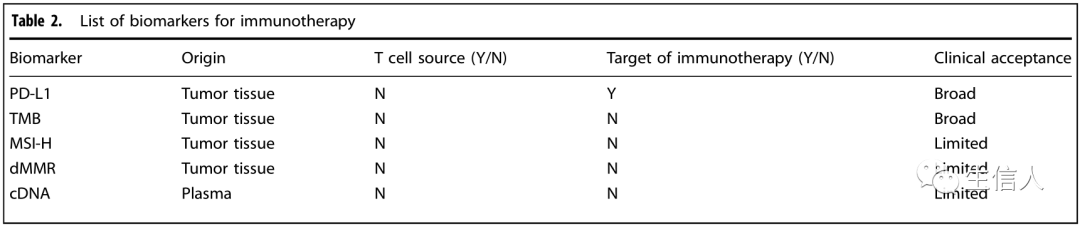
PD-L1被作為抗PD-1治療的第一個生物標志物,被列入pembrolizumab的處方指南中。目前,與免疫療法療效相關的常見或潛在的生物標志物,根據其可及性,主要有以下幾類: (i)?表面標志物, 包括PD-L1和其他一些抑制性受體,可以通過腫瘤組織的免疫組化檢查。 (ii)基因生物標志物, 如腫瘤突變負荷(TMB)、錯配修復系統缺陷(dMMR)、高微衛星不穩定性(MSI-H)、新抗原和抗原呈遞途徑的突變,這些都需要對腫瘤進行基因組分析; ?(iii)?循環腫瘤DNA(ctDNA), 可通過分析外周血獲得。這些生物標志物中的一些已經被III期臨床試驗所驗證,并在臨床上廣泛使用(表2) 。
到目前為止,沒有生物標記物被用于指導患者,但一些CAR-T細胞的內在和外在因素顯示出與治療反應有趣的相關性。腫瘤抗原的表達。CAR-T細胞作為一種靶向治療,需要在腫瘤細胞上表達TAA來誘導T細胞活性,這確實是CAR-T細胞療效最關鍵的生物標志物。由于腫瘤抗原逃逸是CAR-T細胞治療后腫瘤復發的主要機制,也有研究發現,下調TAA不是完全丟失,而是抑制CAR-T細胞功能。雖然CAR-T細胞可以被設計成增加其對低水平TAA的敏感性,但TAA的表達密度可能成為一種預測性的生物標志物。
其他類型的免疫療法
可以預防某些癌癥的發展,包括預防宮頸癌的HPV疫苗、陰道癌、外陰癌、肛門癌和尖銳濕疣以及預防肝癌的HBV疫苗。治療性腫瘤疫苗包括以自由肽或載于APC上的肽的形式注射腫瘤抗原,以激活免疫細胞,恢復其自主抗腫瘤能力。在臨床前模型中,治療性腫瘤疫苗已被證實可以防止癌癥生長和轉移,并減少終止其他類型治療后的復發。 腫瘤疫苗主要分為以下四種:腫瘤全細胞疫苗、基因工程疫苗、蛋白肽疫苗和樹突狀細胞疫苗。
新抗原是存在于癌細胞上的蛋白質片段,提供了一種實現癌細胞特異性靶向的新方法。新抗原疫苗是根據患者的特定腫瘤情況定制的。 生產新抗原疫苗的一般步驟包括(1) 腫瘤活檢,即從患者身上提取腫瘤樣本進行基因組純化;?(2) 對腫瘤細胞和正常細胞的全外顯子組測序,使研究人員能夠尋找腫瘤細胞中獨特的突變;(3) 特異性新抗原的預測和篩選;?(4) 開發個性化疫苗,這是基于預測的新抗原,可以通過多種方法實現,包括多肽、mRNA和DC。最關鍵和最具挑戰性的一步是患者特異性新抗原的鑒定。
有額外病毒感染的癌癥患者通常病情惡化。然而,病毒也可以被修改為專門針對癌細胞。這些“溶瘤病毒”是通過基因組編輯和大規模篩選產生的,其解讀包括對癌細胞的裂解能力,同時保留正常細胞。由此產生的溶瘤病毒抗原可以復制并隨后溶解腫瘤細胞,從而向腫瘤部位釋放更多的病毒顆粒。因此,小劑量的病毒可以在體內擴增。溶瘤病毒具有介導腫瘤抗原擴散的能力,可導致浸潤腫瘤的淋巴細胞增多,增強了ICB治療的抗腫瘤療效。另一種方法是使用溶瘤病毒作為載體,結合細胞免疫治療。通過額外的基因工程,病毒的細胞溶解功能可以被抑制,同時允許合成分子的表達。
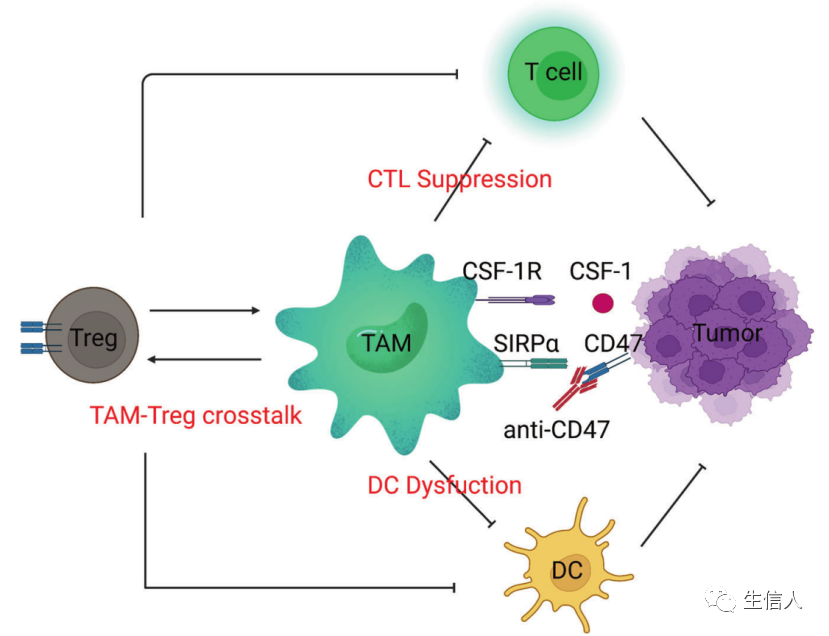
這些先天免疫系統的“士兵”清除了受損、衰老和危險的細胞,但在癌癥中,巨噬細胞促進了它們的免疫逃逸,已成為藥物開發的一個重要領域。 巨噬細胞有多種功能,包括清除細胞碎片和病原體,調節炎癥反應。巨噬細胞也是高度可塑性的細胞,可以根據微環境刺激和信號從一種表型轉換到另一種表型。巨噬細胞的激活狀態通常分為兩類:M1型巨噬細胞和M2型巨噬細胞(圖8)。某些M2巨噬細胞亞群參與促進腫瘤進展和介導免疫抑制。從機制上講,已發現腫瘤將單核細胞和巨噬細胞招募到TME,并將它們極化為M2樣表型。巨噬細胞靶向癌癥治療的中心目標是將腫瘤相關巨噬細胞(TAMs)重編程為促炎(抗腫瘤)亞型。
最成熟的針對TAMs的方法是阻斷集落刺激因子-1(CSF-1,也稱為M-CSF)/CSF1R軸。這種方法減少了TAMs的數量,這也可能與TAMs向M1表型的再極化有關。TAMs的致瘤功能也可以由TGF-β介導,TGF-β是一種抗炎分子,通常在損傷修復過程中由巨噬細胞表達。在一個小鼠模型中,阻斷TGF-β并同時用STING激動劑治療,通過上調I型干擾素的表達而導致腫瘤的消退。Toll樣受體(TLRs)參與先天免疫感應。TLR激動劑可以增加單核細胞的招募/浸潤,并誘導巨噬細胞向促炎癥表型重新極化。TAMs也表達CD40,CD40激動劑可以防止腫瘤生長和減弱抗藥性。
這些抑制性(CSF1R、TGF-β)和刺激性(TLRs、CD40)分子都可以用來恢復TAMs的促炎癥功能。
CD47又稱整合素相關蛋白,屬于免疫球蛋白超家族,通過與巨噬細胞或樹突狀細胞表面的信號調節蛋白α (SIRPα)結合,調節細胞增殖、遷移和凋亡。CD47在大多數腫瘤細胞表面過表達。阻斷CD47/SIRPα通路可誘導巨噬細胞靶向腫瘤的吞噬功能,這在小鼠異種移植模型中得到證實。
巨噬細胞治療的發展需要增加靶向激活受體和更持久的M1巨噬細胞極化方法
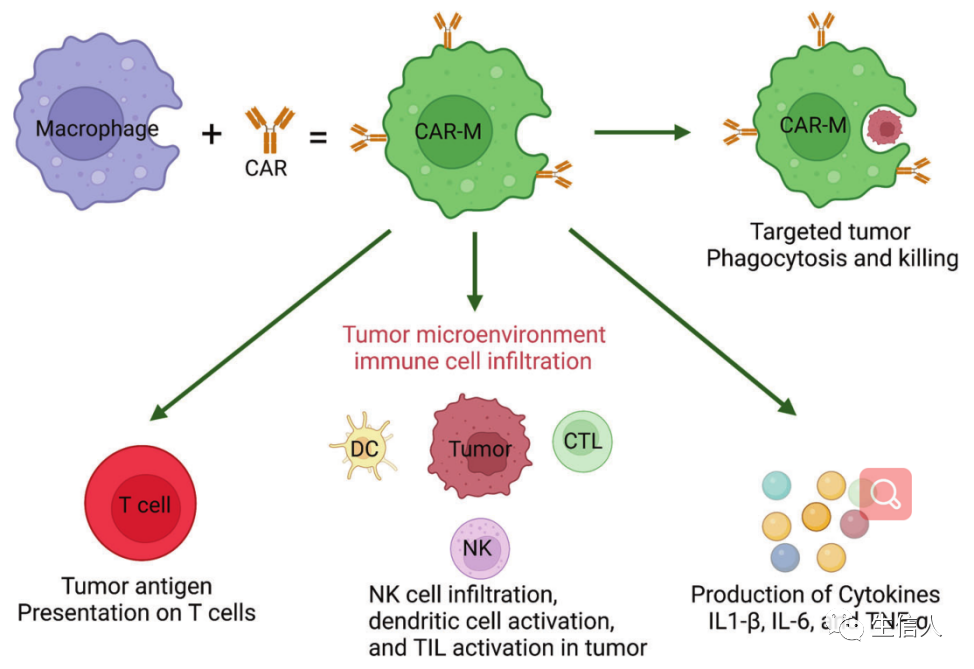
。利用基因修飾增強巨噬細胞的抗腫瘤能力逐漸引起了人們的關注。一個直接的策略是耗盡抑制信號,如SIRPα。SIRPα缺失的巨噬細胞在聯合放療中具有抗腫瘤作用。另一種方法是設計巨噬細胞表達CARs (CAR-Ms)(圖9)。CAR-Ms還誘導周圍TME的促炎特征。M2巨噬細胞的存在不影響CAR-M細胞的腫瘤殺傷能力,突出了其對TME免疫抑制的抵抗力。此外,CAR-Ms表現出更強的T細胞刺
激能力,能夠在吞噬后向T細胞呈遞抗原,招募靜止和激活的T細胞到腫瘤。
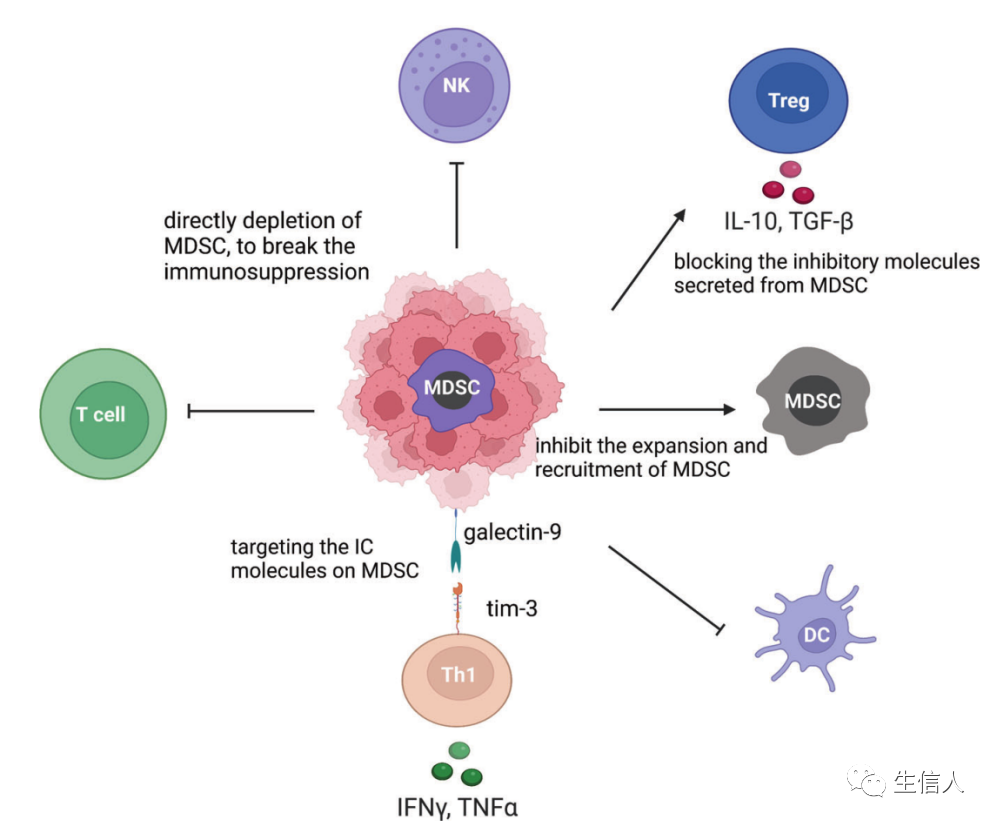
MDSCs起源于造血干細胞(HSCs),是骨髓造血功能改變的結果。這種短暫的骨髓增生在刺激物被移除后會終止,骨髓細胞的平衡也會隨之恢復。然而,在慢性炎癥、癌癥和自身免疫性疾病中,持續的骨髓造血可能發生,以防止宿主廣泛的組織損傷,不斷產生IMCs。這些細胞具有明顯的特征,如不成熟的表型和形態,相對較弱的吞噬功能,以及抗炎和免疫抑制功能。異常的骨髓細胞對其他免疫細胞有抑制作用。此后,人們采用表面標志物Gr-1和CD11b來定義這些免疫抑制性骨髓細胞。在人類中,這些骨髓細胞的表型特征是CD34、CD14和CD15的表達,功能特征是它們能夠抑制T細胞的激活。MDSCs是指一組異質細胞,可大致分為粒細胞(G-MDSCs或PMN-MDSCs)和單細胞(M-MDSCs)亞型。MDSCs的存在是腫瘤進展的基本特征之一。
MDSCs發揮多種功能,影響T細胞、Treg細胞、DC細胞和NK細胞(圖10)。
目前,MDSC靶向治療大致可分為五種類型:(i) 抑制MDSCs擴張和招募的療法;(ii) 恢復正常骨髓分化的療法;(iii) 針對MDSCs上IC分子的療法;(iv) 阻斷MDSCs分泌的抑制性分子的療法;(v) 直接耗盡MDSCs的療法。
腫瘤相關B細胞(TABs)。腫瘤環境中的B細胞(命名為TABs)同時具有促炎和抗炎功能,抗炎B細胞的豐富程度與黑色素瘤對ICB治療的耐藥性相關。TAB在腫瘤組織中的第三淋巴組織結構(TLSs)中富集,在那里它們可以被激活來識別癌細胞。腫瘤中TLSs中B細胞水平高的NSCLC患者更有可能對免疫治療產生良好反應。
IDO 是一種色氨酸代謝酶,可將色氨酸轉化為犬尿氨酸,并在多種類型的腫瘤中過度表達。腫瘤抑制基因BIN1對IDO的表達有負向調節作用。在小鼠模型中,BIN1的耗盡已被證實能誘導IDO的表達和對腫瘤的免疫抑制。IDO可以增強癌細胞的運動能力,抑制腫瘤靶向T細胞的增殖和功能。IDO靶向藥物在臨床前模型中作為單藥顯示出療效,但與靶向CTLA-4或PD-1/PD-L1的ICB聯合使用可以更有效。目前主要有四種針對IDO的小分子抑制劑正在進行臨床研究:indoximod、navoximod、epacadostat和BMS-986205。 IL-41 可增強腫瘤的侵襲性,抑制抗腫瘤免疫。靶向IL-41可能是一種新的免疫治療途徑。 腺苷(Adenosine) 是RNA合成的重要組成部分。然而,腺苷也被證明可以抑制T細胞在TME中的功能。CD39 是一種參與細胞外腺苷產生的酶,在各種人類腫瘤中高度表達。此外,TME中CD39表達水平升高的多種細胞類型包括血管內皮細胞、成纖維細胞和一些免疫細胞。CD39已被發現在多種免疫細胞中發揮重要作用。因此,CD39聯合ICB可能成為一種新的腫瘤免疫治療方式。
這篇文章介紹了腫瘤免疫治療的歷史,深入探討了癌癥免疫治療的不同策略,包括IC和刺激性分子靶向藥物、細胞免疫治療和抑制性TME靶向策略。此外,作者還討論了與ICB和CAR-T治療的臨床療效相關的生物標志物。總之,腫瘤免疫學研究與免疫細胞的先進技術相結合,將為癌癥免疫治療的未來發展指明方向。癌癥免疫治療的發展需要更多的臨床和基礎研究項目的整合,從而能夠全面分析未滿足的臨床需求,進而指導研究方向。 參考文獻:
Wang DR, Wu XL, Sun YL. Therapeutic targets and biomarkers of tumor immunotherapy: response versus non-response. Signal Transduct Target Ther. 2022 Sep 19;7(1):331. doi: 10.1038/s41392-022-01136-2. PMID: 36123348; PMCID: PMC9485144.
文章來源:生信人? ?撰稿:Jennifer,本文由醫世象 佐羅整編編輯。
編輯:小果果,轉載請注明出處:http://www.448371.com/cells/myxb/27547.html
免責聲明:本站所轉載文章來源于其他平臺,主要目的在于分享行業相關知識,傳遞當前最新資訊。圖片、文章版權均屬于原作者所有,如有侵權,請及時告知,我們會在24小時內刪除相關信息。
說明:本站所發布的案例均摘錄于文獻,僅用于科普干細胞與再生醫學相關知識,不作為醫療建議。






 微信掃一掃
微信掃一掃  支付寶掃一掃
支付寶掃一掃 