

介紹
盡管嵌合抗原受體(CAR)T細胞在治療血液系統惡性腫瘤方面已顯示出顯著成果(1),但針對實體癌的有效CAR T治療方法的開發仍然是一項挑戰,很大程度上是由于難以確定最佳目標表面抗原。極少的抗原是真正的腫瘤特異性抗原,與正常組織的靶點/腫瘤外交叉反應會導致致命的毒性(2-5)。此外,即使鑒定出高度腫瘤特異性的抗原,這些靶標也常常是異質表達的,選擇性CAR靶向可以使抗原陰性的腫瘤細胞逃逸(6)。因此,通常需要新穎的腫瘤識別策略,其可以應對特異性和異質性的雙重挑戰,以擴大用于安全有效地攻擊實體癌的治療窗口。
在膠質母細胞瘤(GBM)中發現了這種雙重挑戰的具體例子(圖1A)。表皮生長因子(EGFRvIII)是一種高GBM特異性新抗原,存在于一部分GBM患者中(7-10)。但是在先前的臨床研究中,以EGFRvIII CAR靶向GBM導致了腫瘤復發,盡管有效殺死了EGFRvIII+細胞,但EGFRvIII表達的高度異質性卻使EGFRvIII-腫瘤細胞得以逃脫(6、11、12)。相反,替代的神經膠質瘤相關表面抗原,包括Aphrin A型受體2(EphA2)和IL13受體α2(IL13Rα2),在絕大多數GBM細胞中表達(13-15),但特異性不完善。盡管它們在正常腦組織中不表達,但在一些非腫瘤,非腦組織中卻以低水平表達(13-15)。總而言之,找到單一的,既特異性又均質的理想表面GBM抗原具有挑戰性。
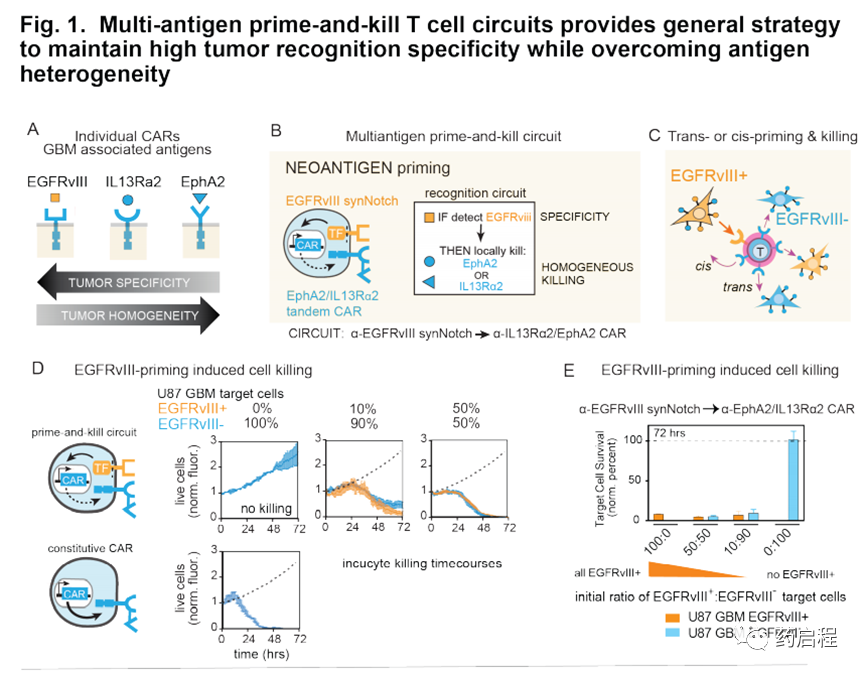

互補的抗原識別回路
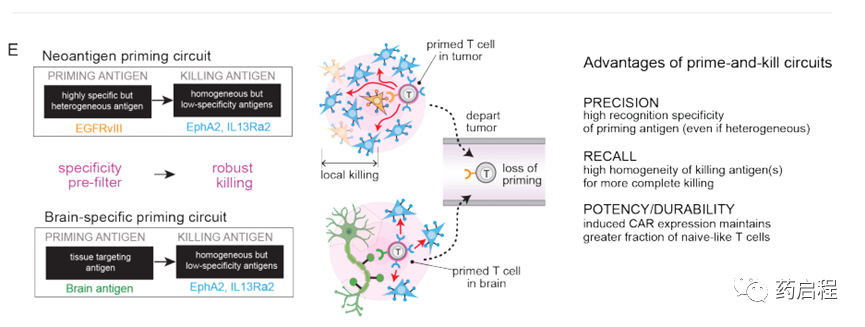
識別多種抗原組合的T細胞為抗原異質性和特異性問題提供了可能的解決方案。我們先前開發了“啟動和殺死”回路,其中synNotch受體(一種工程受體,當其識別其同源抗原時會激活轉錄輸出(16),誘導針對殺傷抗原的CAR的表達(17、18)。在這里我們假設,通過仔細選擇引發和殺死抗原(分別是synNotch和CAR配體),這樣的回路可能導致雜交識別行為,這可能提供一種在抗原特異性和異質性之間進行權衡的方法。
我們采用兩種不同的策略-用腫瘤特異性但異質抗原(例如EGFRvIII)引發(圖1B-E)或用腦特異性抗原(例如髓鞘少突膠質糖蛋白(MOG))引發(圖1F-H )。這些抗原引發的啟動子隨后被用于局部誘導識別更均一的抗原EphA2和IL13Rα2的CAR的表達。EphA2和IL13Rα2的腫瘤特異性不完善,使其成為常規單靶CAR T細胞治療方法的非理想靶標。
但是,如果通過啟動抗原提供更高的腫瘤選擇性,這些抗原可以作為有效的殺傷靶標。我們假設,用引發和殺死回路工程化的T細胞可以誘導局部CAR驅動的細胞毒性,僅局限于引發細胞附近,從而避免了在表達殺傷抗原但缺乏引發抗原的遠距離正常組織中的隨意殺傷。這種類型的回路在空間上整合了對兩個不完美但互補的抗原靶標的識別:啟動抗原提供特異性,而殺傷抗原則確保治療攻擊的均一性。在所有這些回路中,為了實現均勻殺傷并進一步降低腫瘤逃逸的可能性,我們利用了同時靶向兩種殺傷抗原EphA2或IL13Rα2的串聯CAR(13、19)。串聯CAR用作OR門,其胞外區域包含一個α-EphA2單鏈抗體和一個IL13突變蛋白(IL13配體的變體,與IL13Rα1相比與IL13α2的親和力更高)(19、20)。
為了克服這種異質性,一個關鍵問題是由一個細胞(EGFRvIII+)引發的致敏和殺傷性T細胞是否可以殺死另一個相鄰的靶細胞(EGFRvIII-)–我們稱其為反引發過程/殺死(圖1C)。
為了進行轉殺的首次測試,我們使用了經過工程設計以穩定表達啟動抗原EGFRvIII的U87“啟動”GBM細胞和天然U87“靶標”細胞(內源性表達EphA2和IL13Rα2殺傷抗原,但對EGFRvIII陰性)(11,12,21,22)。我們以不同的比例混合了U87-EGFRvIII+和U87-EGFRvIII-細胞,以概括在GBM患者(10-100%的啟動細胞)中觀察到的不同水平的異質性,然后測試了啟動細胞的存在是否誘導了靶細胞的殺傷(圖1D,E)。
我們發現,用α-EGFRvIIIsynNotchàαIL13Rα2/ EphA2 CAR引發和殺死回路改造的CD8+ T細胞可以在體外有效殺死EGFRvIII-靶細胞,即使低至10%EGFRvIII+引發細胞也是如此(圖1D,E)。相反,在不存在引發細胞的情況下,未觀察到對EGFRvIII-靶細胞的殺傷。在這些分析中,我們追蹤了72小時內殺死兩種不同腫瘤細胞群體(EGFRvIII-和EGFRvIII+)的動力學。用低至10%的啟動細胞觀察到有效殺傷,盡管與用50%的啟動細胞觀察到的殺傷相比稍慢(p=.0149;t檢驗)。我們還驗證了用模型抗原進行反式殺傷的有效性,從而顯示了反式啟動/殺傷的耐用性。所有這些體外殺傷研究均表明,有效地進行了啟動引發/殺死,并且EGFRvIII引發的回路因此代表了一種防止因異質性導致的腫瘤逃逸的有前途的策略。
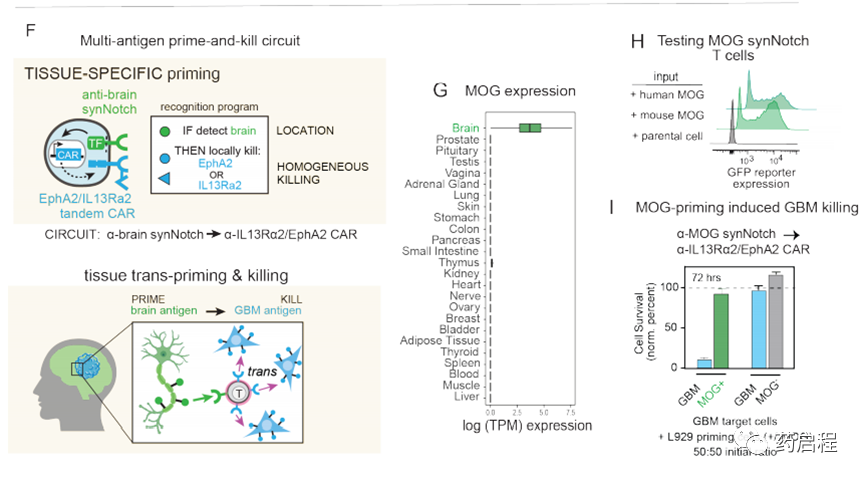
我們還假設,T細胞也可以通過識別組織特異性抗原(在非惡性細胞上表達)來局部引發(圖1F)。例如,在GBM的情況下,我們可能設計由腦特異性抗原引發的T細胞回路,然后通過誘導針對GBM抗原EphA2和IL-13Rα2的CAR來觸發局部殺傷。因此,腦抗原引發的回路可能提供治療EGFRvIII陰性GBM腫瘤的解決方案。我們通過生物信息學鑒定了兩種候選的大腦表面蛋白,Cadherin 10(CDH10)–一種大腦特異性的鈣粘著蛋白,以及髓磷脂少突膠質細胞糖蛋白(MOG)–一種神經元髓鞘上的表面蛋白。這些抗原的預測組織表達如圖1G和圖S7A所示。
在本研究中,我們主要專注于使用MOG作為啟動抗原,因為事實證明它具有更強的大腦特異性(圖S7A中顯示了CDH10作為啟動抗原的分析)。我們鑒定了與MOG結合的抗體,并用它們構建了同源的synNotch受體,這些受體可以被表達MOG小鼠同工型的細胞激活(圖1H),從而使這些受體可由內源性小鼠腦組織引發。然后,我們使用α-MOGsynNotchàαIL13Rα2/ EphA2 CAR回路對CD8+ T細胞進行工程改造,并在存在或不存在引發細胞的情況下將它們與GBM目標細胞(此處為GBM6 PDX細胞系)共培養(經改造的L929細胞表示MOG)。我們發現這些T細胞可以有效殺死GBM細胞,但只有在MOG +引發細胞存在的情況下(圖1I)。重要的是,這些T細胞沒有顯示出對啟動細胞的任何殺死。總而言之,這些體外研究表明,存在多種設計用于針對GBM的引發和殺死回路的策略,這些策略執行反引發/殺死,因此有可能克服抗原異質性,同時仍保持高特異性。
基于這些體外數據,我們接下來在GBM異種移植小鼠模型中評估了這些引發和殺死CAR T細胞的抗腫瘤活性。首先,我們想確認由EGFRvIII引發的T細胞在體內也可以對EGFRvIII-GBM細胞進行轉殺,但僅在存在EGFRvIII+引發細胞的情況下。作為原理上的證明,我們向NCG小鼠植入了雙重腫瘤–在大腦中,我們植入了U87腫瘤,其比例為50%EGFRvIII+和50%EGFRvIII-;在側面,我們植入了U87腫瘤(僅EGFRvIII)(圖2A)。在此,側腹腫瘤代表潛在的交叉反應性正常 表達殺傷抗原但不表達啟動抗原的組織;相反,腦腫瘤既具有啟動抗原又具有殺死抗原。腫瘤接種后第6天,小鼠接受靜脈內注射。給予初免和殺傷性CAR T細胞或對照未轉導的T細胞(n=6 /組)。所有用對照T細胞治療的小鼠均在兩個部位均顯示出腫瘤生長,并迅速達到安樂死終點,中位生存期為25.5天。相比之下,用原始殺傷性CAR T細胞治療的小鼠與對照小鼠相比,表現出對顱內腫瘤生長的顯著抑制(p ?t檢驗)。然而,重要的是,與對照組相比,用致敏和殺傷性CAR T細胞治療的小鼠對側腹腫瘤沒有統計學上的顯著抑制(p = 0.4;t檢驗,圖2B)。在非引發性側翼腫瘤中選擇性缺乏殺傷作用表明,引發性和殺傷性CAR T細胞的細胞毒性活性在空間上局限于表達引發性和殺傷性抗原的腫瘤。
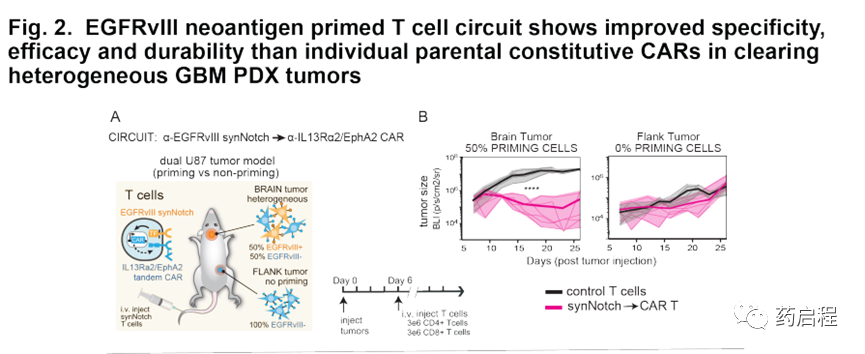
我們還對0%,50%和100%EGFRvIII+ U87細胞殺死植入的腫瘤進行了系統比較(圖S3 A,B)。我們發現,致敏和殺傷性CAR T細胞未顯示出0%EGFRvIII陽性腫瘤的任何清除率,但是顯示了50%和100%EGFRvIII陽性腫瘤的等效清除率(對照未顯示清除率)。因此,在這種情況下,引發和殺死CAR T細胞可以識別并有效克服具有異源EGFRvIII表達的腫瘤,但是可以以特異性的引發抗原門控方式進行。
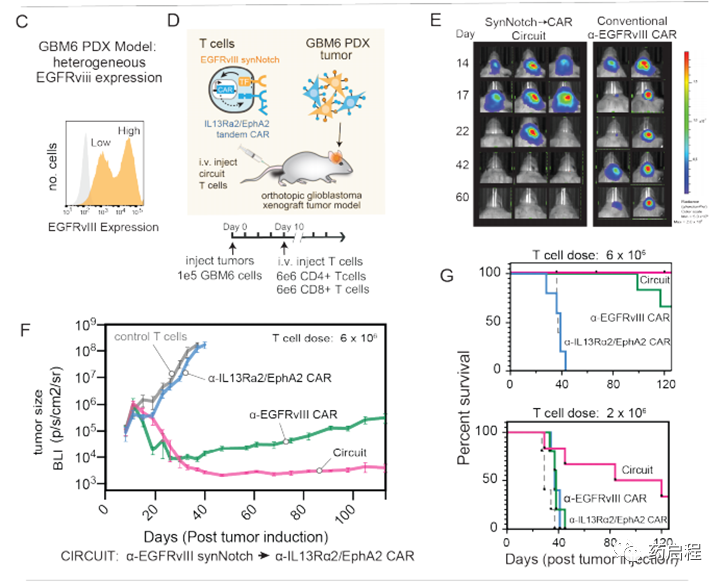
然后,我們試圖評估在腫瘤模型中引發和殺死CAR T細胞的功效,該模型表現出EGFRvIII表達的天然異質性。我們將GBM6患者來源的異種移植(PDX)腫瘤鑒定為具有侵略性的GBM模型,顯示出內在的EGFRvIII異質性(23)(圖2C)。同樣重要的是,GBM6腫瘤在體內逃避EGFRvIII單抗原CAR的治療表現出高度可復制的能力(圖2F)。當用α-EGFRvIIICAR治療小鼠時,GBM6腫瘤急劇縮小,但隨后又緩慢而穩定地復發,并具有很高的再現性(圖2F,圖S3C,如下所述)。這些復發性腫瘤表現出EGFRvIII表達的喪失(圖2J)(我們在體外證實存在一部分GBM6細胞,這些細胞顯示出無法檢測到的EGFRvIII抗原,并且對常規EGFRvIII CAR的殺傷具有抵抗力-圖S4 C,D)。因此,GBM6腫瘤模仿了在α-EGFRvIIICAR臨床試驗中觀察到的基于異質性的逃逸,因此代表了一種理想的腫瘤模型,可以在其中評估可以克服這些問題的替代回路。
我們測試了帶有EGFRvIII引發回路的T細胞,組成型表達α-EGFRvIIICAR或α-IL13Rα2/EphA2串聯CAR的T細胞,非轉導(對照)T細胞對大腦中帶有GBM6腫瘤的NCG小鼠的治療(圖2F,G)。在腫瘤接種后第43天,所有接受非轉導的對照T細胞的小鼠(n = 5)都死于腫瘤進展(圖2E,F)。用組成型α-IL13Rα2/ EphA2串聯CAR治療在很大程度上無效(盡管這些T細胞在體外具有有效的細胞毒性)(圖S4D)。用α-EGFRvIIICAR T細胞治療可產生初始腫瘤縮小,但始終如一(n = 6)導致所有小鼠中EGFRvIII陰性腫瘤復發(6只小鼠中有3只在第125天死于腫瘤進展)(圖2 E,縮略詞)。與之形成鮮明對比的是,所有用原發性和殺傷性CAR T細胞治療的小鼠(n = 6)均顯示GBM6腫瘤的長期完全緩解。這種更持久,更徹底的腫瘤清除率具有很高的重現性(圖S3C),這也反映在用引發和殺傷回路(多次給藥)治療的小鼠的存活率顯著提高的情況下(圖2G)。
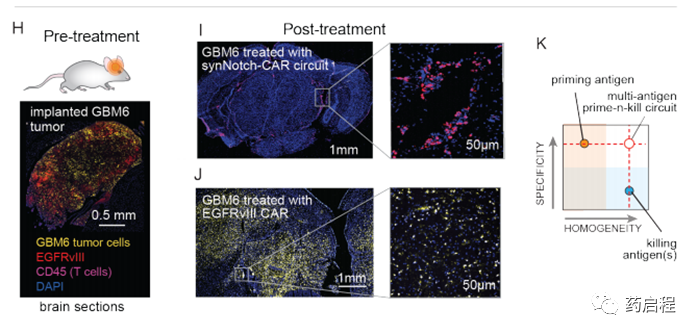
我們對用EGFRvIII CAR或引發殺傷回路處理的小鼠進行了事后免疫熒光分析。用灌注和殺傷回路治療的小鼠的大腦顯示不存在GBM6腫瘤細胞(與腫瘤清除率一致),但在腦實質和腦膜中顯示出CAR T細胞(粉紅色)的持久性(圖2I)。相反,用EGFRvIII CAR治療的大腦顯示出大量的GBM6腫瘤細胞(黃色–與復發一致),但是EGFRvIII抗原丟失(紅色),而CAR T細胞沒有存活(圖2J)。總之,體內腫瘤殺傷研究支持這樣一個概念,即雙抗原回路可以將引發抗原的特異性與殺傷抗原的同質性相結合,以實現比針對單個抗原的CAR所能實現的特異性更高但更徹底的殺傷。
為了直接觀察體內T細胞啟動,我們將GFP標簽融合到了誘導的α-IL13Rα2/EphA2 CAR上。T細胞輸注后六天,對來自受體小鼠的腦切片的分析顯示,腫瘤中存在GFP+引發和殺傷性CAR T細胞(與人CD45共同染色)(圖3A)。相反,在腫瘤外部(鄰近腦組織)或脾臟中均未發現引發的T細胞(圖3A)。此外,我們在注射后兩天對這些致敏和殺死的T細胞進行了活體成像,結果顯示腫瘤中有大量致敏的(綠色)T細胞,以及接近腫瘤時變為綠色的T細胞( 圖3B,電影S2)。重要的是,這些電影表明,引發的T細胞穩定地定位在腫瘤內(大概與靶細胞相互作用),并且沒有迅速進入或移出腫瘤。這種行為可能有助于解釋這些T細胞的高度特異性和局部殺傷作用。
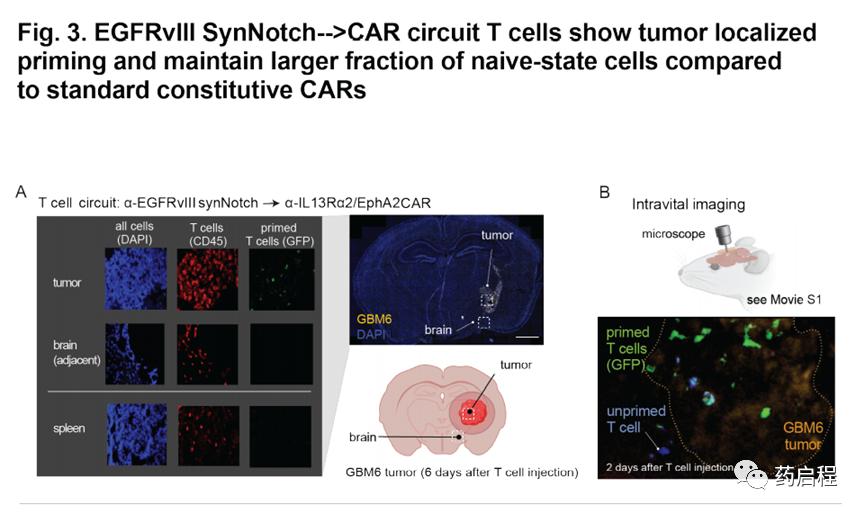
這些體內研究中最令人驚訝的發現之一是,與組成型αIL13Rα2/ EphA2 CAR T細胞相比,初免和殺傷性T細胞的腫瘤清除能力明顯更好,因為兩組T細胞均使用相同的CAR殺死分子,兩者在體外殺死腫瘤方面同樣有效。
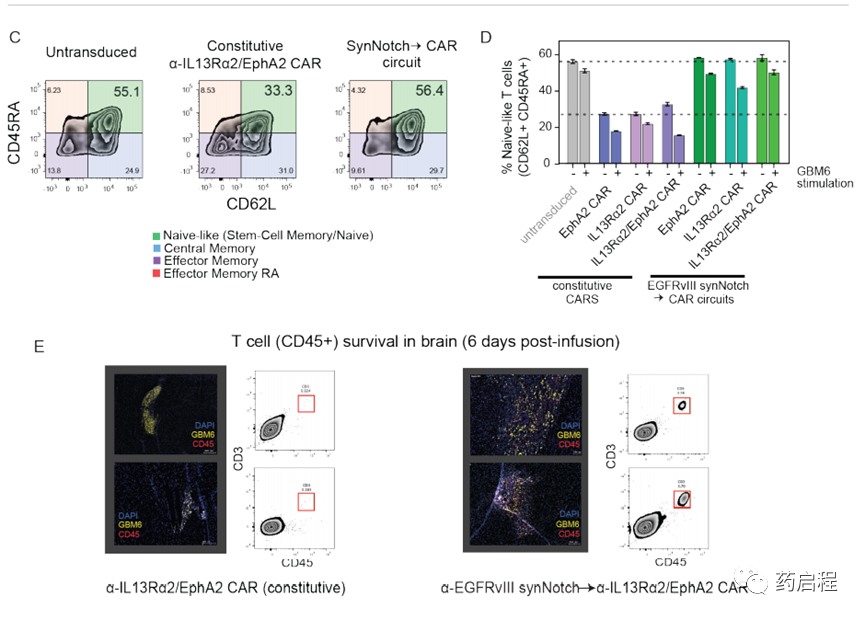
這些觀察結果表明,synNotch誘導的CAR回路具有其他功能,可顯著提高體內的抗腫瘤活性。用CAR T細胞治療實體癌的一般挑戰是T細胞的耗盡,這阻止了持久的抗腫瘤活性。最近的研究表明,組成型表達的CAR的強直信號可以在提高其疲憊敏感性中起重要作用(24,25)。因此,我們通過流動分析檢查了不同類型T細胞的分化狀態,發現本研究中使用的所有synNotchàCART細胞在幼稚樣狀態(CD62L+ CD45RA+,幼稚或干中央記憶)中均顯示出更高比例的細胞。)與等效的組成型CAR T細胞相比(圖3C,D)。此外,當我們直接研究體內T細胞的持久性時(注入小鼠后6天),我們觀察到了大量的synNotchàCAR回路T細胞。相反,我們發現此時沒有存活的組成型串聯CAR T細胞(圖3E)。在一起,這些發現與一個簡單的模型是一致的:synNotchàCAR回路阻止了通常在組成性表達的CAR中觀察到的強直性信號傳導,從而使T細胞保持了更幼稚的狀態,不易耗盡。因此,CAR表達的局部瞬時引發不僅增加了靶向特異性,而且似乎產生了更有效和持久的T細胞狀態。
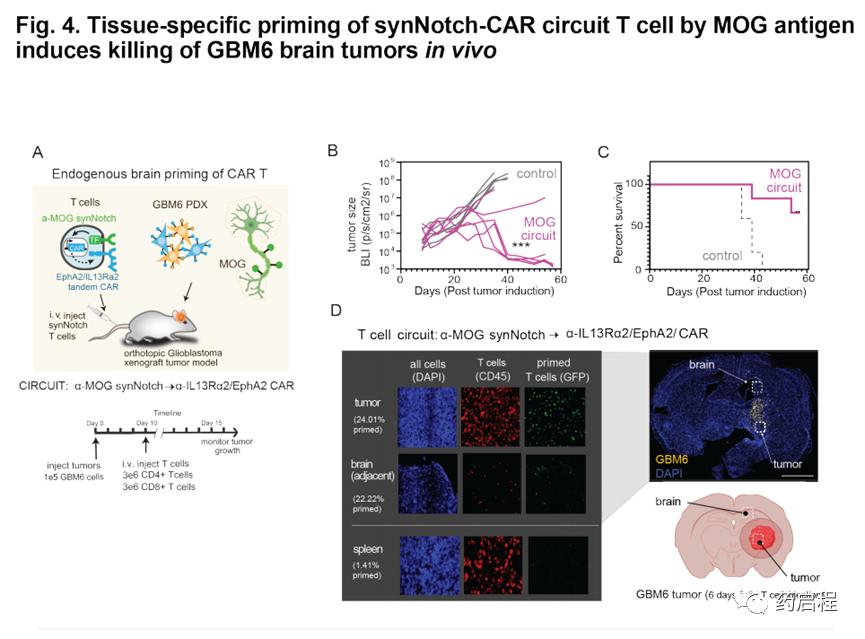
我們還想測試腦特異性抗原引發的T細胞在體內是否有效。因此,我們用植入帶有α-MOGsynNotchàα-IL13Rα2/EphA2 CAR回路的T細胞治療了植入顱內GBM6 PDX腫瘤的NCG小鼠(圖4A)。GBM6腫瘤細胞不表達MOG,因此為了引發T細胞,必須通過宿主小鼠大腦內源性表達的MOG引發它們(圖1I)。我們發現,MOG引發的T細胞在清除GBM6腫瘤和增加小鼠存活率方面非常有效(圖4 B,C)。在將GBM6腫瘤植入側面而不是大腦的小鼠中,MOG引發的T細胞無法清除腫瘤(圖S7F),這與需要局部腦信號以許可T細胞進行殺傷相一致。用α-CDH10synNotchàα-IL13Rα2/EphA2 CAR回路治療的小鼠也顯示出對GBM6的有效殺滅,但在這種情況下,在殺死腦瘤和側翼腫瘤方面顯示出差的辨別力,這表明MOG優于CDH10,因為它嚴格限制了大腦的活動,特異的啟動抗原。
對用MOG引發的T細胞治療的小鼠進行的事后免疫熒光分析顯示,腫瘤中存在大量的T細胞,其中許多處于引發狀態(通過GFP融合CAR觀察)(圖4D)。在腫瘤鄰近的腦組織中,我們觀察到較少的T細胞,但它們也被引發(GFP+)(在脾臟中未觀察到引發的T細胞)。這些結果與整個腦內的T細胞引發,推測與CAR活化和增殖性細胞因子釋放導致的更明顯的T細胞在腫瘤中的擴散相一致。
這種腦特異性抗原引發回路的有效性代表了工程治療細胞的新進展-該回路不僅代表了治療EGFRvIII-GBM的可能方法,而且還可能為工程針對腦的細胞療法提供一般策略 治療廣泛的神經系統疾病,包括其他腦部腫瘤,神經炎癥或神經退行性疾病。
討論
總之,這些結果表明,有多種方法可以設計synNotchàCAR回路,這些回路可以有效地結合識別不完美的互補抗原作為單個抗原靶標。可以建立基于高度特異性的新抗原(如EGFRvIII)或組織特異性抗原(如MOG)引發的回路。至關重要的是,這些引發抗原不需要全部存在于所有腫瘤細胞上,也不必全部存在于任何腫瘤細胞上(在組織特異性引發的情況下)。一旦啟動,就可以對T細胞進行編程,使其靶向腫瘤上同質的CAR抗原,從而完成完整的腫瘤殺死,即使它們自身具有不完善的特異性。通過整合來自多種抗原和多種細胞的信息,這些回路從本質上為我們提供了將腫瘤細化為復雜組織的改進能力,從而為如何以更安全,更具體的方式識別和攻擊腫瘤開辟了許多新的可能性。其他相關策略則將CAR和雙特異性銜接子結合起來以整合多抗原組合(26)。
我們在這里顯示synNotchàCART細胞具有將其與常規CAR T細胞區分開的多個特征,這可能證明在治療實體癌(例如GBM)方面具有很高的優勢。首先,它們的多抗原識別增強了腫瘤細胞與正常細胞之間的區別。其次,這些回路介導反式啟動/殺死的能力使它們能夠克服靶抗原異質性的逃逸。最后,將CAR表達置于受控控制之下的簡單行為似乎將T細胞維持在更加幼稚的狀態,這種狀態更加持久且不易疲勞。對于針對除GBM以外的其他癌癥的synNotchàCAR T細胞,觀察到了類似的改善的抗腫瘤活性(27),這表明這些回路可能為治療許多實體癌提供了非常有效的一般策略。


D
O
i
:
Multi-antigen recognition circuits overcome challenges of specificity, heterogeneity, and durability in T cell therapy for glioblastoma
https://doi.org/10.1101/2021.01.07.425632
編輯:小果果,轉載請注明出處:http://www.448371.com/cells/gxb/9189.html
免責聲明:本站所轉載文章來源于其他平臺,主要目的在于分享行業相關知識,傳遞當前最新資訊。圖片、文章版權均屬于原作者所有,如有侵權,請及時告知,我們會在24小時內刪除相關信息。
說明:本站所發布的案例均摘錄于文獻,僅用于科普干細胞與再生醫學相關知識,不作為醫療建議。
 微信掃一掃
微信掃一掃  支付寶掃一掃
支付寶掃一掃 